
Pyura imesa, Rocha & Counts, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4564.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:FDADF9CA-379C-4980-8AE4-D47F58537E3F |
|
DOI |
https://doi.org/10.5281/zenodo.5930326 |
|
persistent identifier |
https://treatment.plazi.org/id/81CA0AEF-68EF-4C1A-B7A0-F8BDD1CACC15 |
|
taxon LSID |
lsid:zoobank.org:act:81CA0AEF-68EF-4C1A-B7A0-F8BDD1CACC15 |
|
treatment provided by |
Plazi |
|
scientific name |
Pyura imesa |
| status |
sp. nov. |
Pyura imesa View in CoL sp. nov.
( Figures 3 View FIGURE 3 , 4 View FIGURE 4 )
urn:lsid:zoobank.org:act:81CA0AEF-68EF-4C1A-B7A0-F8BDD1CACC15
Pyura lignosa: Tokioka, 1972: 403 View in CoL , fig. 10, 65–70;? Van Name, 1945: 336.
Not Pyura (Halocynthia) lignosa Michaelsen, 1908: 256 View in CoL .
Materials examined: Holotype: MZUSP554 View Materials Isla Canales de Tierra , Shark Point, Pacific coast of Panama, 7°44'18”N 81°34'46.8”W, leg. R. M. Rocha, 10.01.2009. GoogleMaps
Paratype: DZUP PYU-126 Isla Canales de Tierra , Shark Point, Pacific coast of Panama, 7°44'18”N 81°34'46.8”W, leg. R. M. Rocha, 10.01.2009 GoogleMaps .
Etymology. The name is a homage to the Integrated Molecular Ecology of Sponges and Ascidians lab at The University of North Carolina Wilmington for their contributions to ascidian molecular ecology.
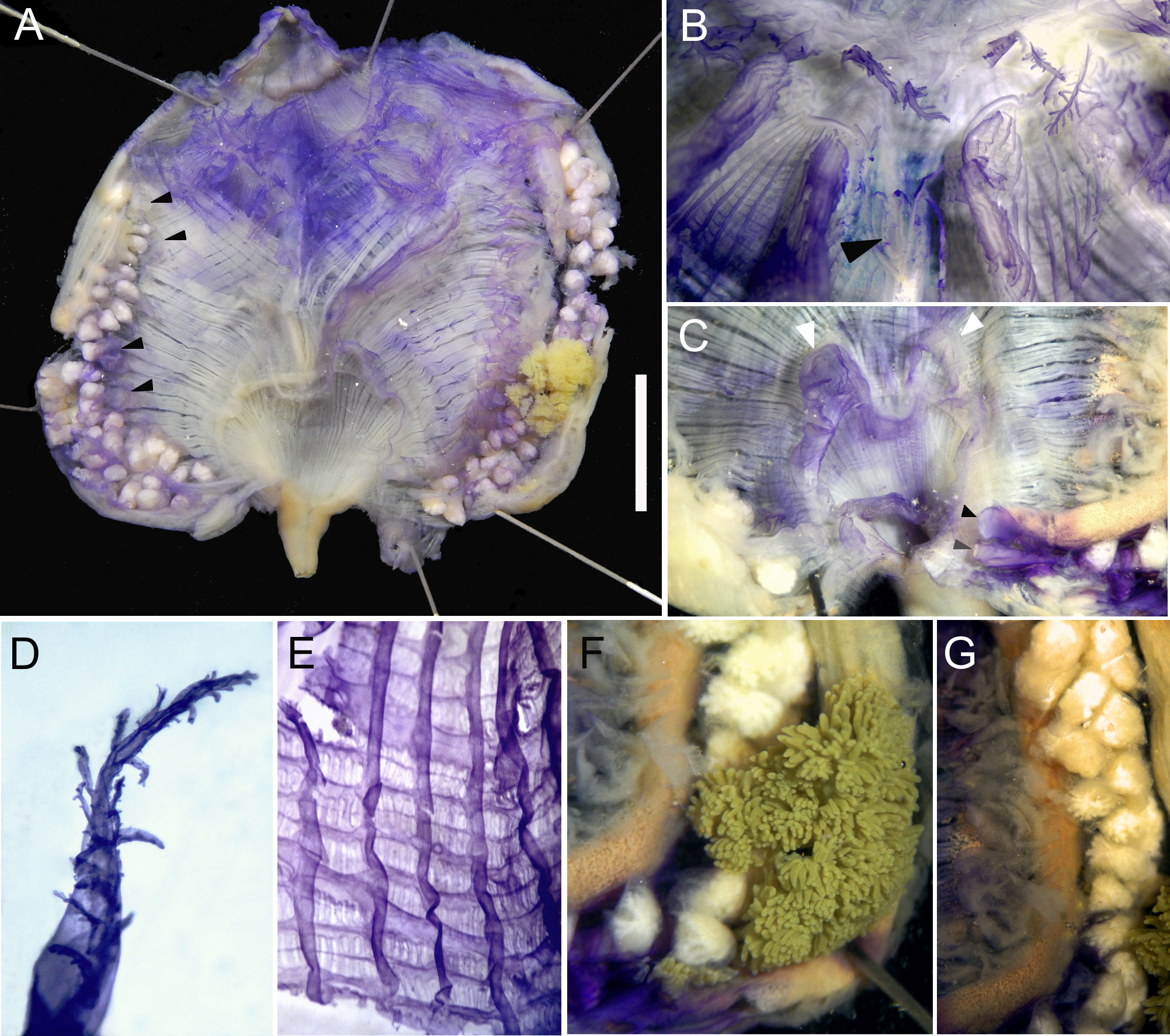
Description. Only two specimens of this species were found in the collection. They were found under rocks, attached by their left or posterior region. Living specimens are orange even in preserved samples ( Fig. 3A View FIGURE 3 ), and pearly white on the inside. The tunic is tough, 1–2 mm thick, and divided into small patches on the surface. The siphons are distant from each other with many tubercles around them, but no spinules or scales on their rim or internal lining. The holotype is 4 cm at its longest length and it is 2 cm high including the tunic, and it is slightly laterally flat. The siphons are both 0.9 cm long without the tunic.
The body wall is cream colored after fixation. Thin circular muscle fibers are densely packed around the siphons. Longitudinal muscles radiate from the siphons and are placed so tightly together in wide bands that they form a continuous sheet, resulting in the body wall being opaque in the anterior half of each side ( Fig. 3B, C View FIGURE 3 ). There is a large velum in the oral siphon.
The tentacles are at the level of a strong band of circular muscles. There are 16 total in the holotype, the largest are more prevalent, with just a few small tentacles occasionally in-between. The right side of the specimen presents more tentacles than the left side where the tentacles are very spaced (1.0 mm apart from each other). The paratype has 12 oral tentacles. They are short, laterally flattened, with slightly large base. They have long second order ramifications lined along the middle of each side or displaced posteriorly, and minute second order ramifications ( Fig. 4B, D View FIGURE 4 ). A small distance, 0.05 mm, separates the tentacles and the prepharyngeal groove which is made up of two equal size lamellae. The lamellae form a deep V around the distinctive V-shaped dorsal tubercle ( Fig. 4B View FIGURE 4 ).
The dorsal lamina does not start immediately after the prepharyngeal groove; instead, it starts after a short distance and it is divided in 90 languets (holotype) that are often found facing toward the right side. The length of the languets increases posteriorly and each languet lines up with the transverse vessels of the pharynx. The pharynx presents six folds per side. The vessel formula of the holotype is (from right to left, for a total of 200):
E 3 (10) 3 (14) 2 (15) 2 (16) 3 (13) 3 (16) 3 DL 2 (13) 2 (14) 2 (15) 2 (14) 2 (14) 2 (12) 3 E
Parastigmatic vessels are frequent but not in all meshes and there are usually 5 or 6 stigmata per mesh between folds and 4 or 5 on folds ( Fig. 4E View FIGURE 4 ).
The intestine is isodiametric with a long narrow primary loop that occupies ¾ of the ventral mid-line of the body and without a secondary loop, ending in a bilabiated smooth anus ( Fig. 4A View FIGURE 4 ). The atrial siphon base is very large with a wide and waving velum with the two dorsal waves larger than the others, but no filiform projections ( Fig. 4C View FIGURE 4 ). The digestive gland is small, yellow (or green in the paratype) and formed by a main portion of tubular projections with two main connections to the stomach, and a few more small portions along the esophagus ( Fig. 4F View FIGURE 4 ).
There are endocarps on the body wall along outer side of the gut, the ones between the ascending intestine and the endostyle are laminar while the ones close to the descending limb are more dense, fleshy and folded ( Fig. 4G View FIGURE 4 ). There are no endocarps attached to the body wall inside the intestinal loop. There are also a few endocarps on the body wall lining the dorsal margin of the right gonad, the two most anterior very small, and the two half the way down, larger and folded ( Fig. 4A View FIGURE 4 ). The paratype have the two small anterior endocarps only. Endocarps are also found on the top of some of the gonadal lobes.
The left gonad lies in the primary intestinal loop, occupying all the space. The gonad lobes are irregularly shaped but tend to be elongated and positioned in pairs along the duct ( Fig. 4A, G View FIGURE 4 ). The right gonad is ventrally placed close to the endostyle and it is longer than the left gonad. There are 37 lobes on the right side and 30 lobes on the left side in the holotype, and 33 on the right and 24 lobes of the left in the paratype. Male follicles tend to ramify when well developed. The gonoducts are equally short, wide and lobed ( Fig. 4C View FIGURE 4 ).
Remarks. Tokioka (1972) described three animals from Playas del Coco, Costa Rica, under the name P. lignosa , but we believe that those animals belong in P. imesa . Although smaller, his exemplars have larger number of oral tentacles and gonadal lobes, and have small spinules around the siphons that we could not find in ours. Otherwise, the tessellated surface of the tunic and presence of tubercles around the siphons, ventral position of the gonads and disposition of endocarps in the body wall agree with our samples. The small animals described by Van Name (1945) under the name P. lignosa and collected in Gulf of California also seem to belong to P. imesa , extending its distribution to the whole Pacific coast of Central America and Mexico. As noted by Monniot (1994), this species is similar to P. vannamei , but we believe that the lack of large endocarps in the right side of the body wall, lack of dilated hind gut and lobes at the margin of the anus, and the geographical distribution justifies naming the Pacific population as a new species. Differences from P. beta Skinner, Rocha & Counts, 2019 are the absence of a second intestinal loop, absence of endocarps inside the primary loop, smaller size and, accordingly, smaller number of oral tentacles, longitudinal vessels and gonadal lobes in P. imesa ( Skinner et al. 2019) . Pyura imesa lives underneath rocks similarly to P. carmanae , however, the color and shape of the animal (distance between siphons), as well as the gonad shape, endocarp abundance and position, distinguish those species.

| DZUP |
Universidade Federal do Parana, Colecao de Entomologia Pe. Jesus Santiago Moure |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Tunicata |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pyura imesa
| Rocha, Rosana M. & Counts, Bailey Keegan 2019 |
Pyura lignosa: Tokioka, 1972 : 403
| Tokioka, T. 1972: 403 |
| Van Name, W. G. 1945: 336 |
Pyura (Halocynthia) lignosa
| Michaelsen, W. 1908: 256 |