
Aleochara (Coprochara) verna Say, 1833
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3641.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:8E36BE56-61EA-4FB9-BEF8-BCDA043EA6DE |
|
DOI |
https://doi.org/10.5281/zenodo.6149269 |
|
persistent identifier |
https://treatment.plazi.org/id/03D94D1B-FFD7-FFFB-53C4-FF41C08F8F47 |
|
treatment provided by |
Plazi |
|
scientific name |
Aleochara (Coprochara) verna Say, 1833 |
| status |
|
Aleochara (Coprochara) verna Say, 1833 View in CoL
Figs. 1 View FIGURES 1 – 4 , 5, 7 View FIGURES 5 – 13 , 14, 21 View FIGURES 14 – 21 , 42 View FIGURES 42 – 43 , 44 View FIGURES 44 – 45
Aleochara (Coprochara) verna ; Say, 1833: 58 (original description); Klimaszewski, 1984: 22; Lohse, 1986: 95; Lohse, 1989: 239; Welch, 1997: 3, 14; Maus, 1998: 87, 96; Maus et al., 1998: 241; Maus & Ashe, 1998 (online); Naomi et al., 2000: 107; Maus et al., 2001: 205; Smetana, 2004: 356; Gouix & Klimaszewski, 2007: 25; Park et al., 2011.
Aleochara languida Sachse, 1852: 117 .
Baryodma pauxilla Mulsant & Rey, 1874: 159 .
Aleochara alticola Sharp, 1883: 148 .
Baryodma minuta Casey, 1906: 161 . [homonym].
Baryodma pumilio Casey, 1911: 6 .
Baryodma tolerata Casey, 1911: 6 .
Aleochara tecumsehi Muona, 1977: 16 . [replacement name].
Aleochara tanumi Likovský, 1984: 8 . [replacement name].
Aleochara cedari Likovský, 1984: 8 . [replacement name].
“ Aleochara bipustulata ”; Sharp, 1888: 282 (as Aleochara nitida Gravenhorst , a synonym of A. bipustulata ); Adachi, 1957: 34; Naomi, 1989: 280; Smetana, 2004: 355.
See Klimaszewski (1984) and Gouix & Klimaszewski (2007) for further references.
Materials examined. JAPAN: [Hokkaidô]: 1 3, Memuro-dake (Penkenûshi-dake), Shimizu-chô, Kamikawa-gun, 15. VII. 1995, K. Haga leg. (cHaga); 1 3, 1 Ƥ, Shari-dake, Shari-machi, Shari-gun, 23. VII. 1996, K. Mizota leg. (KUM). [Honshû]: 1 3, Matsugabô-dam, Yamakami, Sôma-shi, Fukushima-ken, 20. XII. 2006, S. Saitô leg. (leaf litter in forest; cWat); 1 Ƥ, Azuma-yama, Inawashiro-machi, Yama-gun, Fukushima-ken, 19. VIII. 1996, S.-I. Naomi leg. (KUM); 2 3, Ômomo, Minamiaizu-machi, Aizu-gun, Fukushima-ken, 23. VI. 1990, T. Watanabe leg. (cWat); 1 3, 2 sex?, Maeshirane-san, Chûgushi, Nikkô-shi, Tochigi-ken, 23. VI. 1990, T. Watanabe leg. (cWat); 4 sex?, Yumoto, Nikkô-shi, Tochigi-ken, 29. VI. 1982, S.-I. Naomi leg. (KUM); 1 sex?, same data, but 10. VII. 1994; 2 sex?, Chûzenji, Nikkô-shi, Tochigi-ken, 28–30. VI. 1982, S.-I. Naomi leg. (KUM); 1 Ƥ, Akagi-san (Komagatake peak), Fujimi-mura, Seta-gun, Gunma-ken, 20. VII. 1996, S. Nomura leg. (KUM); 1 Ƥ, Hôshi-onsen hotspring, Minakami-machi, Tone-gun, Gunma-ken, 26. VI. 1997, S.-I. Naomi leg. (KUM); 2 3, 6 ƤƤ, Akaguna-yama, Kamihino, Fujioka-shi, Gunma-ken, 13. IX. 2001, T. Watanabe leg. (cWat); 1 3, same data, but 15. V. 2002; 1 3, Watarase-yûsuichi retarding basin, Fujioka-shi, Tochigi-ken, 13. IV. 2002, T. Watanabe leg. (cWat); 3 3, 3 ƤƤ, Matsuzawa-tôge pass (alt. 1150m), Haruna-san, Takasaki-shi, Gunma-ken, 27. VII. 1996, S. Nomura leg. (KUM); 3 3, Nidoage-tôge pass, Kawaura, Kurabuchi-machi, Takasaki-shi, Gunma-ken, 11. IX. 2002, T. Watanabe leg. (cWat); 6 sex?, Kanaibuchi-machi, Takasaki-shi, Gunma-ken, 27. III. 2005, T. Watanabe leg. (cWat); 1 3, 7 sex?, Wagamine-machi, Takasaki-shi, Gunma-ken, 27. III. 2005, T. Watanabe leg. (cWat); 1 3, Kirizumi, Sakamoto, Matsuida-machi, Annaka-shi, Gunma-ken, 19. VIII. 1962, Y. Shibata leg. (cShi); 3 3, 3 ƤƤ, Kamioshidari, Higashimatsuyama-shi, Saitama-ken, 1. XI. 1998, K. Toyoda leg. (KUM); 1 3, Tenmoku-zan, Chichibu-shi, Saitama-ken, 12. VI. 1998, K. Toyoda leg. (KUM); 3 ƤƤ, Hirogawarasawa, Nakatsugawa, Chichibu-shi, Saitama-ken, 11. X. 1999, K. Toyoda leg. (KUM); 1 Ƥ, Tone-gawa, Sakae-machi, Inba-gun, Chibaken, 5. V. 1991, T. Takeda leg. (KUM); 1 Ƥ, Amatsu-kaigan, Uchiura, Kamogawa-shi, Chiba-ken, 3. XI. 1990, T. Takeda leg. (KUM); 1 Ƥ, Heisaura, Sakai, Tateyama-shi, Chiba-ken, 1. IV. 1997, M. Maruyama leg. (KUM); 1 sex?, river mouth of Obitsu-gawa, Kuroto, Kisarazu-shi, Chiba-ken, 25. X. 1998, K. Toyoda leg. (KUM); 1 sex?, Takao-san, Hachiôji-shi, Tôkyô-to, 16. IV. 1962, Y. Shibata leg. (cShi); 1 Ƥ, Arasaki, Nagai, Yokosuka-shi, Kanagawa-ken, 17. XI. 1963, Y. Shibata leg. (cShi); 1 sex?, Hayama-machi, Miura-gun, Kanagawa-ken, 27. III. 1953, R. Kiryû leg. (SCM); 1 sex?, Kamimizo, Chuô-ku, Sagamihara-shi, Kanagawa-ken, 23. V. 1954, R. Kiryû leg. (SCM); 1 3, Shônan-kaigan, Kanagawa-ken, 2. VII. 1976, Niizato leg. (without collector’s given name; KUM); 1 Ƥ, Mitsuishi (Manazuru-misaki), Manazuru-machi, Ashigarashimo-gun, Kanagawa-ken, 15. VII. 1989, T. Watanabe leg. (under rubbish on the beach; cWat); 1 Ƥ, Susugaya (Tanzawayamafudakake), Kiyokawa-mura, Aikô-gun, Kanagawa-ken, 6. XI. 1993, T. Watanabe leg. (cWat); 1 3, 1 sex?, Tônosawa, Hakone-machi, Ashigarashimo-gun, kanagawa-ken, 24. X. 1971, Y. Hirano leg. (KUM); 1 Ƥ, Usui-tôge, Karuizawa, Karuizawamachi, Kitasaku-gun, Nagano-ken, 31. VII. 1999, S.-I. Naomi leg. (KUM); 13 sex?, Kamihikawa, Daibosatsu, Kôshû-shi, Yamanashi-ken, 14. VI. 1986, T. Watanabe leg. (cWat); 4 3, 1 Ƥ, near Tsuchimuro-gawa, Daibosatsu, Yamanashi-ken, 16–17. X. 1982, Y. Shibata leg. (cShi); 1 sex?, Hôoh-goya lodge, Minamiarupusu-shi, Yamanashiken, 26. VII. 1967, Y. Shibata leg. (cShi); 1 Ƥ, same locality, 11. VI. 1990, K. Hosoda leg. (cIto); 1 Ƥ, same data, but 2. VI. 1993; 1 sex?, Hirogawara (Shirane-san), Minamiarupusu-shi, Yamanashi-ken, 9–13. VII. 1982, S.-I. Naomi leg. (KUM); 1 3, 2 sex?, Yanbushi-tôge pass, Hayakawa-chô, Minamikoma-gun, Yamanashi-ken, 4. VII. 2002, A. Sugimura leg. (KUM); 1 Ƥ, 1 sex?, river mouth of Yodo-gawa, Ôsaka-shi, Ôsaka-fu, 17. III. 1956, Y. Hayashi leg. (cHay). [Shikoku]: 1 3, 2 ƤƤ, 1 sex?, Ôtaki-san, Shionoe-chô, Takamatsu-shi, Kagawa-ken, 8. VI. 2008, H. Fujimoto leg. (KUM); 2 3, Tsurugi-san, Tokushima-ken, 19–20. VI. 1981, S.-I. Naomi leg. (KUM). [Kyûshû]: 1 3, 19 sex?, Shakaga-dake (alt. 1200m), Yame-shi, Fukuoka-ken, 25. IX. 1994, S. Nomura leg. (KUM); 2 sex?, Sobo-san, Ôita-ken, 19. VII. 1985, K. Ogata leg. (KUM); 1 3, 2 ƤƤ, 1 sex?, Sobo-san, Taketa-shi, Ôita-ken, 7. VI. 2009, S. Yamamoto leg. (daytime on walkway; cYam); 13 sex?, same data, but 14. VII. 2011 (besides these records, dozens of individuals were observed (only field observation, no collecting permit) at the top of the mountain which is situated on a boundary of Ôita-ken and Miyazaki-ken: Fig. 42 View FIGURES 42 – 43 ); 3 ƤƤ, 2 sex?, Kurokawa, Aso-shi, Kumamoto-ken, 6. VI. 2009, S. Yamamoto leg. (from cow dung on pasture during daytime; cYam); 1 3, 1 Ƥ, Ichifusa-yama, Mizukami-mura, Kuma-gun, Kumamoto-ken, 1–2. VIII. 1988, S.-I. Naomi leg. (KUM); 1 Ƥ, Tomioka (Amakusa-syotô), Reihoku-machi, Amakusa-gun, Kumamoto-ken, 3. IV. 1977, S.-I. Naomi leg. (KUM); 1 3, Nichinan-shi, Miyazaki-ken, 29. V. 1969, Watanabe leg. (without collector’s given name; KUM); 1 Ƥ, Wanitsuka-yama, Kou, Tano-chô, Miyazaki-shi, Miyazaki-ken, 6. IX. 1993, S. Nomura leg. (KUM); 1 sex?, Kirishima-yama, Kobayashi-shi, Miyazaki-ken, 4. VII. 1982, M. Takanashi leg. (KUM); 1 3, Takachiho, Miyazaki-ken, 24. VI. 1969, Y. Goto leg. (cShi); 28 sex?, Takachihonomine, Miike-chô, Miyakonojô-shi, Miyazakiken, 21. V. 1982, H. Takemoto leg. (KUM); 1 sex?, same locality, 19. VII. 1983, M. Ôhara leg. (KUM); 8 sex?, Takachihonomine, 31. V. 1981, Y. Takai leg. (KUM); 5 sex?, same locality,? VIII. 1979, Koda leg. (without detail collecting date and collector’s given name; KUM); 1 Ƥ, 19 sex?, Takachihonomine, kirishimataguchi, Kirishima-shi, Kagoshima-ken, N. Koda leg. (without collecting date; KUM); 1 3, 1 Ƥ, 3 sex?, Kirishima-yama (near Miike pond), Kagoshima-ken, 18. V. 1983, Y. Sawada leg. (KUM); 1 3, Unzen-dake, Unzen-shi, Nagasaki-ken, 17. V. 1986, T. Itô leg. (cIto); 3 sex?, same locality, 3. VII. 1977, S. Imasaka leg. (KUM); 1 Ƥ, same data, but 4. VII. 1978; 1 3, 1 sex?, same data, but 24. VI. 1979; 1 Ƥ, same data, but 5. VIII. 1980; 1 Ƥ, same data, but 24. VI. 1981.
Other material. RUSSIA: [Kamchatka Peninsula]: 1 Ƥ, Nagornyi (53°07'N 158°31'E), near Petropavlovsk- Kamachatskiy, 10. VII. 1997, A. Saitô leg. (Naomi et al., 2000; CBM: CBM-ZI/81531).
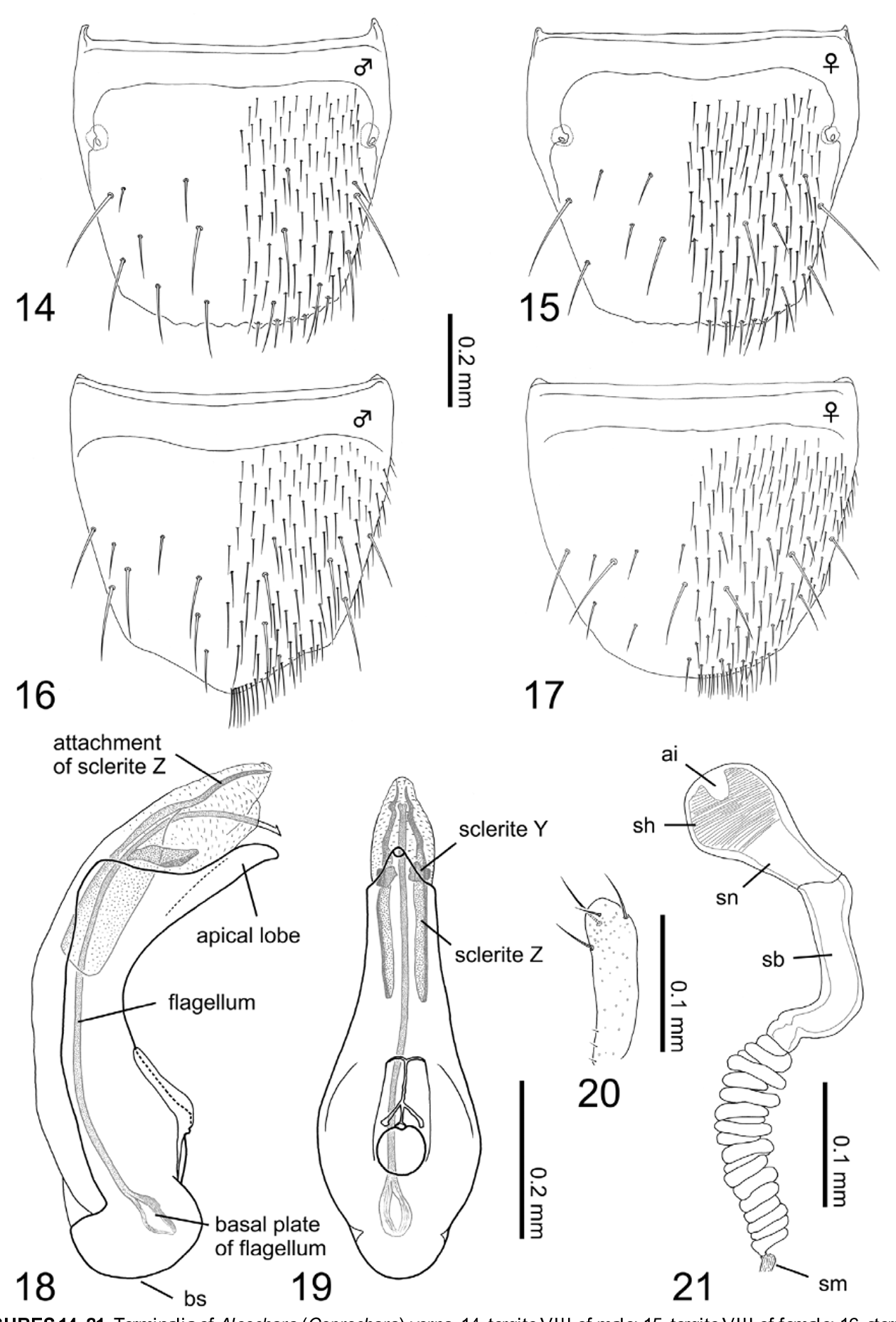
Redescription. Body ( Fig. 1 View FIGURES 1 – 4 ): slender and subparallel, normally moderate in size, ca. 3.63 mm in body length (3.12–4.32 mm, N = 15), and 1.72 mm in fore body length (1.41–1.83 mm, N = 15), robust, and dorsal surface clearly glossy. Color ( Figs. 1 View FIGURES 1 – 4 , 5–7 View FIGURES 5 – 13 ): ground color completely black; legs, especially tarsal segments, and mouth parts brown to reddish brown; elytra with a pair of yellowish spots; antennae blackish brown to reddish brown, surface moderately to densely pubescent with minute whitish setae. Head ( Fig. 1 View FIGURES 1 – 4 ): circular (head length = 0.47– 0.57 mm (mean = 0.53 mm), head width = 0.49–0.63 mm (mean = 0.57 mm), N = 15), slightly transverse (head width / head length = 1.08 (mean), N = 15), widest at eyes; surface covered with long and thin setae sparsely, punctation shallow and inconspicuous. Antennae ( Fig. 1 View FIGURES 1 – 4 ): moniliform, thick and robust, shorter than head and pronotum combined (antenna length = 0.74–0.95 mm (mean = 0.87 mm), N = 15); segment I, about 2.3 times as long as broad, segment II clearly shorter than I, segment III slightly shorter than II, segment IV spherical, as long as width except for stem of segment, segments V to X clearly transverse, segment XI thick and subconical, nearly 1.8 times as long as width, longer than segment I, approximate relative length of segments from basal to apex: 12.0: 10.5: 7.5: 4.0: 4.5: 5.0: 5.0: 5.0: 5.0: 5.0: 15.0. Thorax: pronotum ( Fig. 1 View FIGURES 1 – 4 ) moderately transverse (pronotum length = 0.54–0.79 mm (mean = 0.65 mm), pronotum width = 0.68–0.91 mm (mean = 0.84 mm), pronotum width / pronotum length = 1.29 (mean), N = 15), wider than head (pronotum width / head width =1.49 (mean), N = 15), widest just after middle; surface coarsely and shallowly punctured, with weak longitudinal rows of punctures along midline. Intercoxal process of mesoventrite ( Fig. 7 View FIGURES 5 – 13 ) with strong carina along midline. Intercoxal process of metaventrite ( Figs. 6–7 View FIGURES 5 – 13 ) broad and short, rounded apically. Metaventrite ( Fig. 7 View FIGURES 5 – 13 ) smooth, pubescent like elytra. Elytra ( Fig. 5 View FIGURES 5 – 13 ) wider than long (elytra length = 0.54–0.70 mm (mean = 0.62 mm), elytra width = 0.77–1.11 mm (mean = 0.98 mm), N = 15) with a pair of yellowish small spots extending from posterior margin close to elytral suture; surface shallowly punctured and with inconspicuous setae; posterior margin weakly rounded. Legs ( Fig. 1 View FIGURES 1 – 4 ): short (hind tibia length = 0.44–0.65 mm (mean = 0.55 mm), N =15), dozens of long spines on fore and midtibia, respectively; relative lengths of tarsal segments from basal to apical: 4.5: 3.0: 3.0: 3.0: 8.5 in foretarsus, 9.0: 5.0: 5.0: 5.0: 11.0 in midtarsus, 13.0: 8.0: 8.0: 8.0: 14.5 in hindtarsus. Abdomen ( Fig. 1 View FIGURES 1 – 4 ): weakly glossy, covered by short and thin setae. [ Male]: posterior margin of tergite VIII ( Fig. 14 View FIGURES 14 – 21 ) clearly serrate, with approximately 8 macrosetae on each side. Sternite VIII ( Fig. 16 View FIGURES 14 – 21 ) with approximately 8 macrosetae; posterior margin pointed triangularly. Median lobe of aedeagus as in Figs. 18–19 View FIGURES 14 – 21 : elongated and narrowed toward apex; flagellum inside projecting, longer than median lobe of aedeagus; basal plate of flagellum small and short; sclerite Z with long attachment, waving apically; apical lobe in lateral view moderately bent, narrowing subapically; basal swelling of median lobe circularly rounded; median lobe in ventral view moderately tear-shaped; basal swelling of median lobe small; apical lobe looks short isosceles. Apical lobe of paramerite ( Fig. 20 View FIGURES 14 – 21 ) long and slender, bearing 4 long setae.
[Female]: tergite VIII ( Fig. 15 View FIGURES 14 – 21 ) with weakly serrate posterior margin, with 7 macrosetae. Sternite VIII ( Fig. 17 View FIGURES 14 – 21 ) rounded posteriorly, with 8 macrosetae. Spermatheca ( Fig. 21 View FIGURES 14 – 21 ): apical invagination of spermatheca small but deep; spermathecal head spherical, as long as spermathecal neck; attachment of spermathecal duct inconspicuous; (sn) clearly narrowing toward basal portion of spermathecal stem, stem moderately curved to almost angled at its base; coiled portion of moderate length, composed of maximally 20 thin coils, not strongly extended laterally; coils directly connected with thin membranous portion of spermathecal duct; each part of spermatheca except for (sm) entirely and moderately sclerotized; inner wall of (sh) finely striate.
Diagnosis. This species is similar to the other Japanese species of the subgenus Coprochara , but can be easily distinguished from them by the following character states: surface smooth and glossy ( Fig. 1 View FIGURES 1 – 4 ); elytral posterior margin rounded with a pair of yellowish spots on dorsal surface of elytra ( Figs. 1 View FIGURES 1 – 4 , 5 View FIGURES 5 – 13 ); intercoxal process of metaventrite moderately rounded apically ( Figs. 6–7 View FIGURES 5 – 13 ); tergite VIII without thick spines in posterior half ( Figs. 14– 15 View FIGURES 14 – 21 ). [ Male]: sclerite Z inside the median lobe with long waving attachment at apex ( Figs. 18–19 View FIGURES 14 – 21 ). [Female]: spermatheca with maximally 20 thin coils, not greatly extended laterally ( Fig. 21 View FIGURES 14 – 21 ).
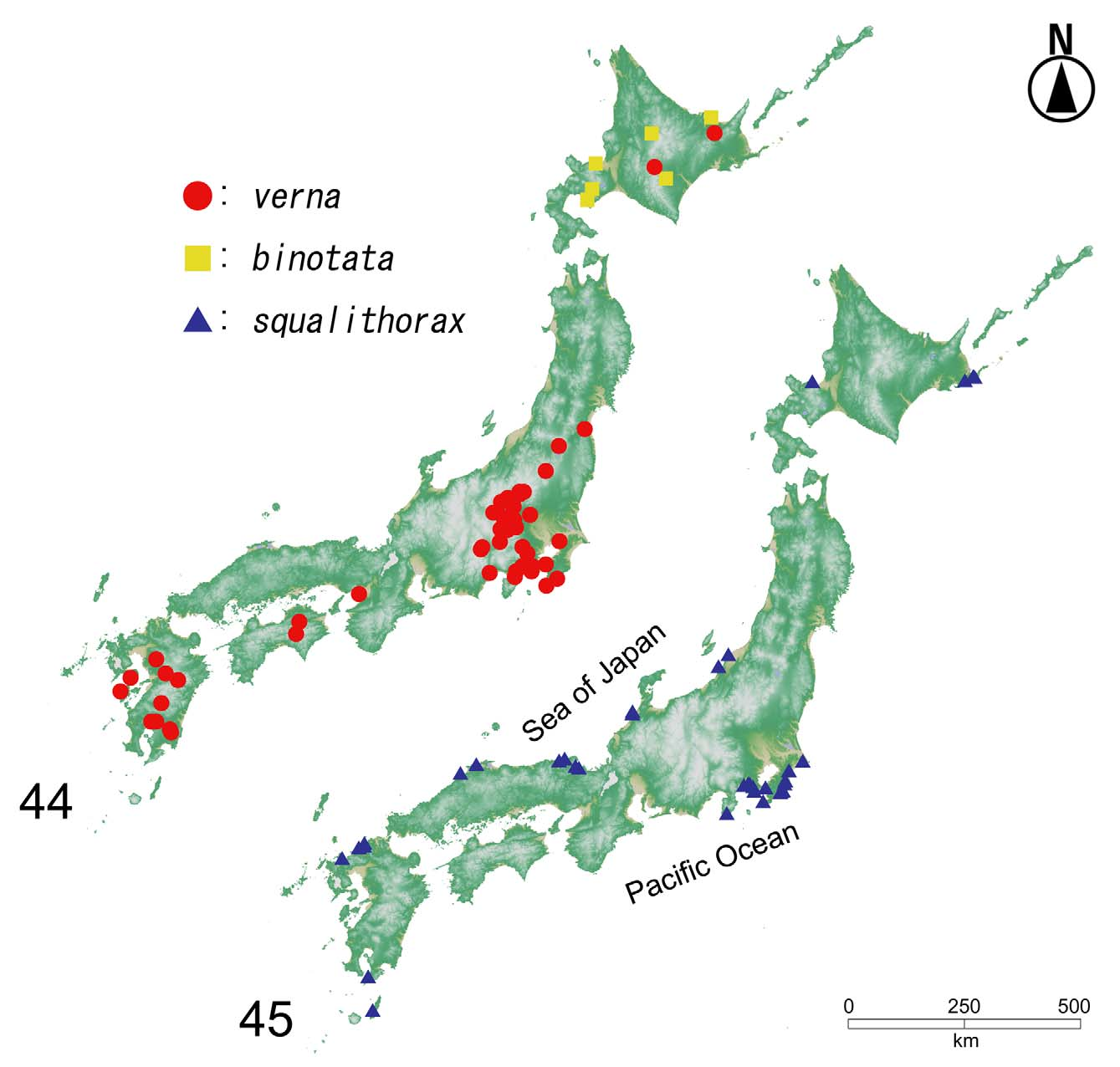
Confirmed distribution by the present study. [ JAPAN]: Hokkaidô, Honshû, Shikoku, Kyûshû, [ RUSSIA]: Kamchatka Peninsula.
Other localities in the literature. [EUROPE]: Austria, Azerbaijan, Azores, Belgium, Bosnia Herzegovina, Bulgaria, Croatia, Czech Republic, Denmark, France, Georgia, Germany, Greece, Hungary, Italy, Lithuania, the Nertherlands, Poland, Portugal, Romania, Russia: South European Territory, Slovakia, Spain, Sweden, Switzerland, UK, Ukraine (Maus, 1998; Smetana, 2004; Assing, 2006; Prado e Castro et al., 2010; Tamutis et al., 2011), [NORTH AFRICA]: Algeria, Canary Islands, Egypt, Libya, Morocco, Madeira Archipelago (Smetana, 2004), [ASIA]: Afghanistan, China, India, Iran, Israel, Kazakhstan, Mongolia, Nepal, North Korea, Russia: Far East, South Korea, Syria, Tajikistan, Turkmenistan, Turkey, Uzbekistan (Maus, 1998; Smetana, 2004; Park et al., 2011), [NORTH AMERICA]: Alaska, Canada, Hawaii, Mexico, USA (Klimaszewski, 1984; Navarrete et al., 2002; Gouix & Klimaszewski, 2007), [SOUTH AMERICA]: Venezuela (Navarrete et al., 2002).
Remarks. Aleochara verna is very common and widely distributed in the Holarctic region. The distribution ranges through all of Europe and North Africa, across northern India and the Russian Far East to North America (Maus, 1998; Smetana, 2004). In Japan, reliable records had been reported from only two localities, Gunma and Kanagawa Prefectures (Maus, 1998; Park et al., 2011). Numerous records are added from all of Japan, from northern Hokkaidô to southern Kyûshû, Japan.
This species was originally described from Missouri, USA. As the type specimen was destroyed, the neotype was designated by Klimaszewski (1984). The distribution of this species in the Palearctic region is unclear; it has been suggested that A. verna is not a Nearctic, but a Palearctic species (Maus, 1998).
The taxonomic status of A. verna was unstable until Lohse (1986) and thus far several cases of confusion have occurred, e.g., misidentifications of species in Europe and the proposed use of many synonyms for some species and subspecies under A. verna (Klimaszewski, 1984; Welch 1997; Maus, 1998). Recent studies such as Welch (1990, 1997) and Maus (1998) have allowed the precise identification of the Coprochara -complex, including A. verna , in the Palearctic region.
The Japanese specimens examined have a blackish body with a pair of small yellow markings on the elytra; no geographical variations were detected. This species is commonly found in Japan; many records have been reported as “ Aleochara bipustulata ”. However, true specimens of A. bipustulata were not found in the present study. Therefore, the distribution of A. bipustulata in Japan is doubtful and it should be excluded from the Japanese fauna (see Discussion).
Park et al. (2011) redescribed and recorded A. verna from South Korea, with diagnoses of A. verna and “ A. bipustulata ”. Following the Japanese example, Korean specimens of “ A. bipustulata ” should be reexamined.
In Japan, Aleochara verna might be the most efficient candidate as a biological control agent due to its abundance, and wide distribution range, and the availability of information that has been accumulated in previous studies on the species.
Bionomics. Species of the subgenus Coprochara occur predominantly in the dung of herbivorous vertebrates and in a wide range of organic materials (Klimaszewski & Jansen, 1994). Aleochara verna is considered as inhabiting in the same environment. Adults were collected from human feces and other animal droppings such as cow, horse, dog, and woodchuck, as well as carrion (Klimaszewski, 1984). The first author collected the species from cow dung in a pasture at Aso, Kumamoto Prefecture, Japan. He also confirmed that A. verna was attracted to small prawns (leavings from a packed lunch) on a walkway in 2009 at Mt. Sobo, Japan. SY also collected specimens of this species on a walkway in summer 2011 on the same mountain, and observed dozens of individuals walking actively on rocks and pebbly surface on the summit ( Fig. 42 View FIGURES 42 – 43 : arrow).
The altitudinal range of A. verna is quite wide. Klimaszewski (1984) noted that adult specimens were collected at altitudes from less than 200 m to nearly 4,000 m. Japanese specimens were found at altitudes from 0 (seashore) to over 2,000 m.
Host records. Comprehensive dipteran host records for A. verna were provided by Maus et al. (1998). This paper gives the following four families as its hosts: Anthomyiidae , Scathophagidae , Muscidae , and Sarcophagidae .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aleocharinae |
|
Genus |
|
|
SubGenus |
Coprochara |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aleocharinae |
|
Genus |
|
|
SubGenus |
Coprochara |