
Aleochara (Coprochara) squalithorax Sharp, 1888
|
publication ID |
https://doi.org/10.11646/zootaxa.3641.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:8E36BE56-61EA-4FB9-BEF8-BCDA043EA6DE |
|
DOI |
https://doi.org/10.5281/zenodo.6149275 |
|
persistent identifier |
https://treatment.plazi.org/id/03D94D1B-FFD0-FFE6-53C4-FE28C6D08FF7 |
|
treatment provided by |
Plazi |
|
scientific name |
Aleochara (Coprochara) squalithorax Sharp, 1888 |
| status |
|
Aleochara (Coprochara) squalithorax Sharp, 1888 View in CoL
Figs. 3, 4 View FIGURES 1 – 4 , 11, 13 View FIGURES 5 – 13 , 30 View FIGURES 30 – 33 , 41 View FIGURES 34 – 41 , 43 View FIGURES 42 – 43 , 45 View FIGURES 44 – 45
Aleochara (Coprochara) squalithorax ; Sharp, 1888: 282 (original description); Adachi, 1957: 34; Naomi, 1989: 280; Assing, 1995: 235; Maus, 1998: 84, 98; Maus & Ashe, 1998 (online); Ahn et al., 2000: 244; Maus et al., 2001: 206; Park & Ahn, 2004: 196; Smetana, 2004: 356; Frank & Ahn, 2011: 19; Yamamoto & Maruyama, 2012: 5, 11.
Type specimens. [Lectotype]: 3, “ Aleochara / squalithorax / Type D.S. / Hakodate / Lewis [PC, HW] // Type [RRL] // Hakodate. // Japan. / G. Lewis // Sharp Coll. / 1905-313 // Lectotypus / design. Assing 1994 // Lecto- / type [PRL]” (abdominal segment VIII-X and aedeagus are dissected and glued on a PC together with the body). Paralectotypes, 1 Ƥ, 2 sex?, “ Aleochara squalithorax / Fauvel. / Hagi. Japan [HW] // SYN- / TYPE [BRL] // Japan. / G. Lewis // Sharp Coll. / 1905-313 // Para- / lecto- / type [BRL]” (three specimens are mounted in the same PC).
Materials examined. JAPAN: [Hokkaidô]: 6 sex? (GT), 2 sex? (YT), Hamamatsu, Nemuro-shi, 14. VI. 2010, S. Yamamoto leg. (from dry seaweed on huge sandy beach; cYam); 2 sex? (GT), Ochiishi, Nemuro-shi, 24. VIII. 1999, M. Maruyama leg. (KUM); 55 sex? (GT), Sanriga-hama, Ochiishi, Nemuro-shi, 12. VI. 2010, S. Yamamoto leg. (from completely dry seaweed on huge sandy beach; cYam); 19 sex? (GT), Urayakotan, Hamanaka-chô, Akkeshi-gun, 12. VI. 2010, S. Yamamoto leg. (from somewhat wet seaweed on sandy beach; cYam); 1 3 (GT), Zenibako, Otaru-shi, 30. V. 1977, N. Nishikawa leg. (KUM). [Honshû]: 4 sex? (GT), Gouya-kaigan, Yotsugouya, Nishi-ku, Niigata-shi, Niigata-ken, 4. V. 1996, K. Haga leg. (under rubbish on sandy beach; cHaga); 13 sex? (GT), Kanayama, Teradomari, Nagaoka-shi, Niigata-ken, 3. V. 2004, M. Nishikawa leg. (KUM); 2 sex? (GT), Uchinadamachi, Kahoku-gun, Ishikawa-ken, 15. VI. 1961, Y. Hayashi leg. (cHay); 1 sex? (GT), Awagasaki, Uchinadamachi, Kahoku-gun, Ishikawa-ken, 12. V. 1948, S. Takaba leg. (cHay); 1 Ƥ (GT), 1 sex? (GT), Hasaki-kaigan, Kamisu-shi, Ibaraki-ken, 13. X. 2009, H. Ono leg. (sandy beach; cOno); 1 3 (GT), 2 ƤƤ (GT), 1 sex? (GT), same data, but 19. X. 2009; 2 3 (GT), 3 ƤƤ (GT), 1 sex? (GT), same data, but 29. X. 2009; 1 3 (GT), 2 ƤƤ (GT), 5 sex? (GT), Ôami, Chiba-ken, 1. V. 1990, M. Ôhara leg. (KUM); 2 3 (GT), 4 3 (YT), 3 ƤƤ (GT), 1Ƥ (YT), 7 sex? (GT), river mouth of Ichinomiya-gawa, Hitotsumatsu, Chôsei-mura, Chôsei-gun, Chiba-ken, 16. V. 1993, T. Takeda leg. (KUM); 2 sex? (GT), Ôhara-kaigan, Ôhara, Isumi-shi, Chiba-ken, 30. V. 1992, H. Oda leg. (KUM); 13 sex? (GT), 2 sex? (YT), Hebara, Katsuura-shi, Chiba-ken, 12. V. 1991, T. Takeda leg. (KUM); 2 3 (YT), 20 sex? (GT), 15 sex? (YT), Katsuura, Katsuura-shi, Chiba-ken, 4. V. 1990, T. Takeda leg. (KUM); 1 Ƥ (YT), 10 sex? (GT), Ubara, Katsuura-shi, Chiba-ken, 6. IV. 1993, T. Takeda leg. (KUM); 1 3 (GT), Heisaura, Sakai, Tateyamashi, Chiba-ken, 13. VIII. 1993, T. Takeda leg. (KUM); 1 3 (YT), 1 Ƥ (GT), 18 sex? (GT), 5 sex? (YT), same locality, but 1. IV. 1997, M. Maruyama leg. (KUM); 1 sex? (GT), Shinmaiko-kaigan, Yawata, Futtsu-shi, Chibaken, 11. VI. 1995, K. Emoto leg. (cWat); 1 sex? (GT), Arasaki, Nagai, Yokosuka-shi, Kanagawa-ken, 12. VI. 2009, T. Watanabe leg. (cWat); 6 sex? (GT), Hayama-machi, Miura-gun, Kanagawa-ken, 28. III. 1954, R. Kiryû leg. (SCM); 1 3 (GT), Yuiga-hama, Kamakura-shi, Kanagawa-ken, 5. IX. 1976, Niizato leg. (without collector’s given name; KUM); 4 sex? (GT), Shichiriga-hama, Kamakura-shi, Kanagawa-ken, 20. IV. 1991, K. Haga leg. (cHaga); 1 sex? (GT), same locality, but 6. V. 1991, Y. Ueda leg. (cWat); 1 3 (GT), Tujidô-kaigan, Tujidô, Fujisawa-shi, Kanagawa-ken, 31. V. 1986, T. Watanabe leg. (under seaweed cast on shore; cWat); 1 Ƥ (GT), Tujidô, Fujisawashi, Kanagawa-ken, 3. V. 1995, T. Watanabe leg. (under seaweed cast on shore; cWat); 14 sex? (GT), Shônankaigan, Kanagawa-ken, 2. VII. 1976, Niizato leg. (without collector’s given name; KUM); 8 sex? (GT), Shirahama, Shirahama, Shimoda-shi, Shizuoka-ken, 13. III. 1989, H. Oda leg. (KUM); 1 3 (GT), Mihama, Maizuru-shi, Kyôto-fu, 3. V. 1980, Y. Hayashi leg. (cHay); 1 sex? (GT), Kanzaki-kaisuiyokujô swimming beach, Nishikanzaki, Maizuru-shi, Kyôto-fu, 2. IV. 2000, M. Yoshii leg. (cHay); 4 sex? (GT), Amino-chô-hamazume, Kyôtango-shi, Kyôto-fu, 21. V. 2000, K. Yasukawa leg. (cHay); 4 sex? (GT), 1 sex? (YT), Kumihama-chô-minatomiya, Kyôtango-shi, Kyôto-fu, 17. IV. 2000, H. Nagata (Osada?) leg. (cHay); 1 3 (GT), Koura-kaigan, Kashima-chô, Matsue-shi, Shimane-ken, 17. VII. 2010, S. Yamamoto leg. (under dry flotsam on sandy beach; cYam); 1 3 (GT), 1 Ƥ (GT), 5 sex? (GT), Taki-chô, Hikawa-gun, Shimane-ken, 2. VI. 1980, S.-I. Naomi leg. (KUM). [Kyûshû]: 5 3 (GT), 1 3 (YT), 2 ƤƤ (GT), 1 Ƥ (YT), 122 sex? (GT), 17 sex? (YT), Watari, Fukutsu-shi, Fukuoka-ken, 21. IV. 2012, S. Yamamoto, M. Maruyama, T. Kanao leg. (from dry to wet seaweed on huge sandy beach; KUM); 5 sex? (GT), 3 sex? (YT), Watari, Fukutsu-shi, Fukuoka-ken, 21. IV. 2012, S. Yamamoto, M. Maruyama, T. Kanao leg. (from dry to wet seaweed on small sandy beach; KUM); 1 3 (YT), 1 Ƥ (YT), 4 sex? (GT), 1 sex? (YT), Mitoma, Higashi-ku, Fukuoka-shi, Fukuoka-ken, 3. VI. 2001, S. Ogata leg. (KUM); 2 sex? (GT), Katsuma (Shikano-shima), Higashi-ku, Fukuoka-shi, Fukuoka-ken, 21. III. 2012, S. Yamamoto leg. (from seaweed on huge sandy beach; cYam); 1 sex? (GT), Karatsu-shi, Saga-ken, 7. VII. 1975, H. Ohishi leg. (KUM); 2 sex? (GT), Minamiôsumi-chô (Sata-misaki), Kimotsuki-gun, Kagoshima-ken, 28. V. 1958, Y. Miyake leg. (cHay); 1 Ƥ (GT), 11 sex? (GT), Nakatane-chô (Tanega-shima), Kumage-gun, Kagoshima-ken, 8-13. V. 1996, M. Maruyama leg. (KUM).
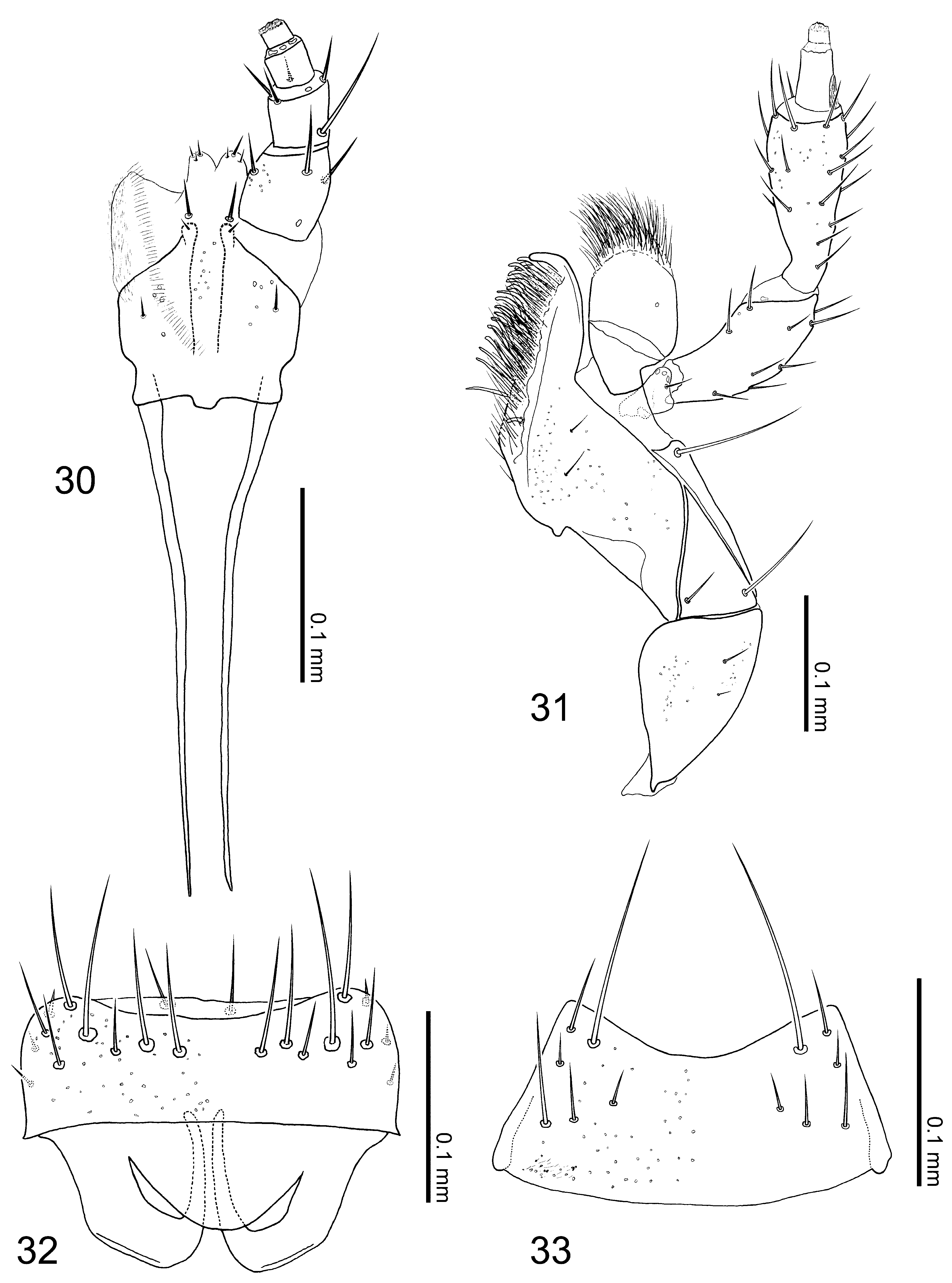
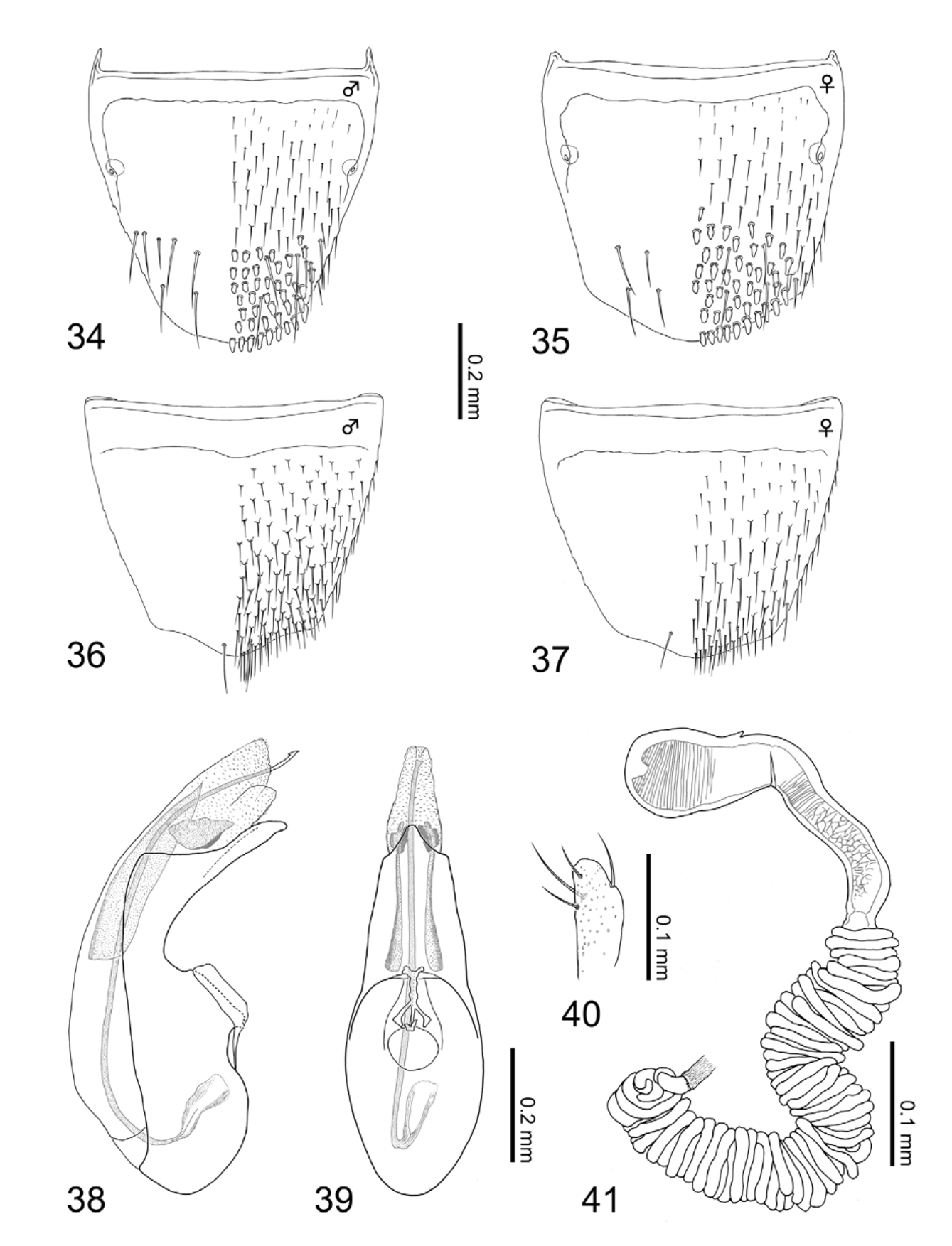
Redescription. Body ( Figs. 3–4 View FIGURES 1 – 4 ): thick and spindle shaped, especially for small individuals ( Fig. 4 View FIGURES 1 – 4 ), small to medium sized, ca. 3.48 mm in body length (3.14–3.83 mm, N = 15), and 1.88 mm in fore body length (1.68–2.15 mm, N = 15), normally moderate in size; entire body heavily robust; dorsal surface mat; forebody covered with granular microstructures. Color ( Figs. 3–4 View FIGURES 1 – 4 , 11–13 View FIGURES 5 – 13 ): ground color gray to blackish brown; legs, especially tarsal segments, and mouth parts dark brown to reddish brown; elytra gray in most cases as Fig. 3 View FIGURES 1 – 4 , but some individuals with huge yellowish portion except for anterior and lateral margins as Fig. 4 View FIGURES 1 – 4 ; antennae reddish brown to dark brown, surface densely pubescent with minute yellowish setae. Head ( Figs. 3–4 View FIGURES 1 – 4 ): circular (head length = 0.42–0.56 mm (mean = 0.48 mm), head width = 0.46–0.56 mm (mean = 0.51 mm), N = 15), slightly transverse (head width / head length = 1.08 (mean), N = 15), widest at eyes; surface entirely covered with hexagonal reticulations, lacking punctation, but covered with short and thick setae sparsely. Antennae ( Figs. 3–4 View FIGURES 1 – 4 ): moniliform, robust and thickened apically, slightly longer than head length (antenna length = 0.57–0.84 mm (mean = 0.73 mm), N = 15); segment I, about 2.1 times as long as broad, segment II slightly shorter than I, segment III apparently shorter than II, segments IV to VI more or less spherical, as long as width except for stem of each segment, segment VII moderately wider than long, segments VIII to IX clearly transverse, segment X strongly transverse, segment XI thick and conical, nearly 1.2 times as long as width, as same length as segment I, approximate relative length of segments from basal to apex: 10.5: 10.0: 6.0: 4.5: 4.5: 4.5: 4.5: 4.5: 4.5: 4.5: 10.5. Mouth parts ( Figs. 30–33 View FIGURES 30 – 33 ): mandibles slightly asymmetric, left one with one tooth near apex. Clypeus rounded apically. Labrum ( Fig. 32 View FIGURES 30 – 33 ) transverse, about 1.3 times as wide as long, anterior margin slightly emarginated medially, basal half semitransparent, apodeme roundly produced postero-medially; surface with numerous pseudopores scattered randomly in apical part. Ligula ( Fig. 30 View FIGURES 30 – 33 ) bilobed, with each lobe rounded apically. Labial palpus ( Fig. 30 View FIGURES 30 – 33 ) with segment I thicker and longer than II, segment II weakly dialated, segment III shorter than II. Mentum ( Fig. 33 View FIGURES 30 – 33 ) nearly trapezoidal, much wider than long, about 2.0 times as wide as long; anterior margin strongly emarginated; entire surface with numerous pseudopores scattered randomly. Maxilla ( Fig. 31 View FIGURES 30 – 33 ): maxillary palpus with thick and long segment II and III, segment II as long as III, segment IV short, less than half length of segment III; galea thick and short, clearly shorter than segment II of maxillary palpus; lacinia with numerous setae and with dozens of thick spines pectinately. Thorax: pronotum ( Figs. 3–4 View FIGURES 1 – 4 ) transverse (pronotum length = 0.60–0.76 mm (mean 0.69 mm), pronotum width = 0.77–0.94 mm (mean = 0.88 mm), pronotum width / pronotum length = 1.27 (mean), N = 15), clearly wider than head (pronotum width / head width = 1.71 (mean), N = 15), widest around basal 1/3, weakly narrowing apically; surface uniformly covered with large hexagonal reticulations, thick-short setae but without distinct punctation; midline longitudinally elevated weakly above, glabrous, lacking punctation. Intercoxal process of mesoventrite ( Fig. 13 View FIGURES 5 – 13 ) with strongly developed carina along midline. Intercoxal process of metaventrite ( Figs. 12–13 View FIGURES 5 – 13 ) narrow and long, apparently pointed apically. Metaventrite ( Fig. 13 View FIGURES 5 – 13 ) rough, densely pubescent like elytra. Elytra ( Figs. 3–4 View FIGURES 1 – 4 , 11 View FIGURES 5 – 13 ) wider than long (elytra length = 0.62–0.78 mm (mean = 0.68 mm), elytra width = 0.90–1.13 mm (mean = 1.04 mm), N = 15), rugose, moderately covered with yellowish thick and short setae; posterior margins deeply notched laterally ( Fig. 11 View FIGURES 5 – 13 ); [Gray type]: dorsal surface entirely gray, but [Yellow type]: with a pair of large yellowish portions, extending from apical to posterior except for anterior margins and shoulders close to pronotum as well as lateral margins of elytra and elytral suture (posterior margins colored). Legs ( Figs. 3–4 View FIGURES 1 – 4 ): short (hind tibia length = 0.49–0.65 mm (mean = 0.57 mm), N = 15) and thick, dozens of thick spines on tibia, especially on fore and midtibia; relative lengths of tarsal segments from basal to apical: 5.0: 3.5: 3.5: 3.0: 8.0 in foretarsus, 8.0: 5.0: 5.0: 4.5: 14.5 in midtarsus, 17.5: 10.0: 7.0: 7.0: 14.5 in hindtarsus. Abdomen ( Figs. 3–4 View FIGURES 1 – 4 ): rogose, except for anterior margin of each tergite and posterior margin of tergite VI to VII; tergite VIII ( Figs. 34–35 View FIGURES 34 – 41 ) with dozens of thick spines scattered in posterior half.
[ Male]: posterior margin of tergite VIII ( Fig. 34 View FIGURES 34 – 41 ) rounded, with approximately 7 macrosetae on each side. Sternite VIII ( Fig. 36 View FIGURES 34 – 41 ) with one macroseta (difficult to identify); posterior margin weakly pointed. Median lobe of aedeagus as in Figs. 38–39 View FIGURES 34 – 41 : compactly elongated and narrowed toward apex; flagellum long, clearly longer than median lobe of aedeagus; sclerite Y large; sclerite Z large without attachment at apex; apical lobe in lateral view slightly bent, narrowing subapically, median lobe in ventral view elongated, weakly narrowing toward apex; basal swelling of median lobe large and oval; apical lobe looks short isosceles. Apical lobe of paramerite ( Fig. 40 View FIGURES 34 – 41 ) short and thick, widest at subapical portion, bearing 4 setae.
[Female]: tergite VIII ( Fig. 35 View FIGURES 34 – 41 ) with rounded posterior margin like male, with approximately 4 macrosetae. Posterior margin of sternite VIII ( Fig. 37 View FIGURES 34 – 41 ) weakly pointed, with one macroseta (difficult to confirm). Spermatheca ( Fig. 41 View FIGURES 34 – 41 ): apical invagination of spermatheca tiny and shallow; spermathecal head fused with spermathecal neck; attachment of spermathecal duct prominent; (sn) short, slightly narrowing toward basal portion of spermathecal stem; coiled portion extremely long, composed with countless times of moderately thick coils, not extended laterally, waving toward membranous portion of spermathecal duct; coils directly connected with thick (sm); each part of spermatheca except for (sm) entirely and strongly sclerotized; inner wall of (sh) coarsely and that of (sb) in apical portion also coarsely striate with rough mesh striate in mid to posterior part of (sb).
Diagnosis. Aleochara squalithorax can be easily distinguished from the other Coprochara in Japan by a combination of the following character states: body thick and spindle shaped; surface mat with granular microstructures, covered with thick and short setae coarsely but without punctation on head and pronotum ( Figs. 3– 4 View FIGURES 1 – 4 ); pronotum clearly wider than head, without rows of punctures along midline; elytra with posterior margin deeply notched, sometimes with yellowish portion on dorsal surface (YT: Figs. 4 View FIGURES 1 – 4 , 11 View FIGURES 5 – 13 ); intercoxal process of metaventrite narrow and sharply pointed apically ( Fig. 13 View FIGURES 5 – 13 ); tergite and sternite VIII with thick spines in posterior half ( Figs. 34– 35 View FIGURES 34 – 41 ). [ Male]: flagellum long; sclerite Z large without attachment ( Figs. 38–39 View FIGURES 34 – 41 ); apical lobe of paramerite short, thick, without numerous minute setae ( Fig. 40 View FIGURES 34 – 41 ). [Female]: countless coiled portion with spermatheca ( Fig. 41 View FIGURES 34 – 41 ).
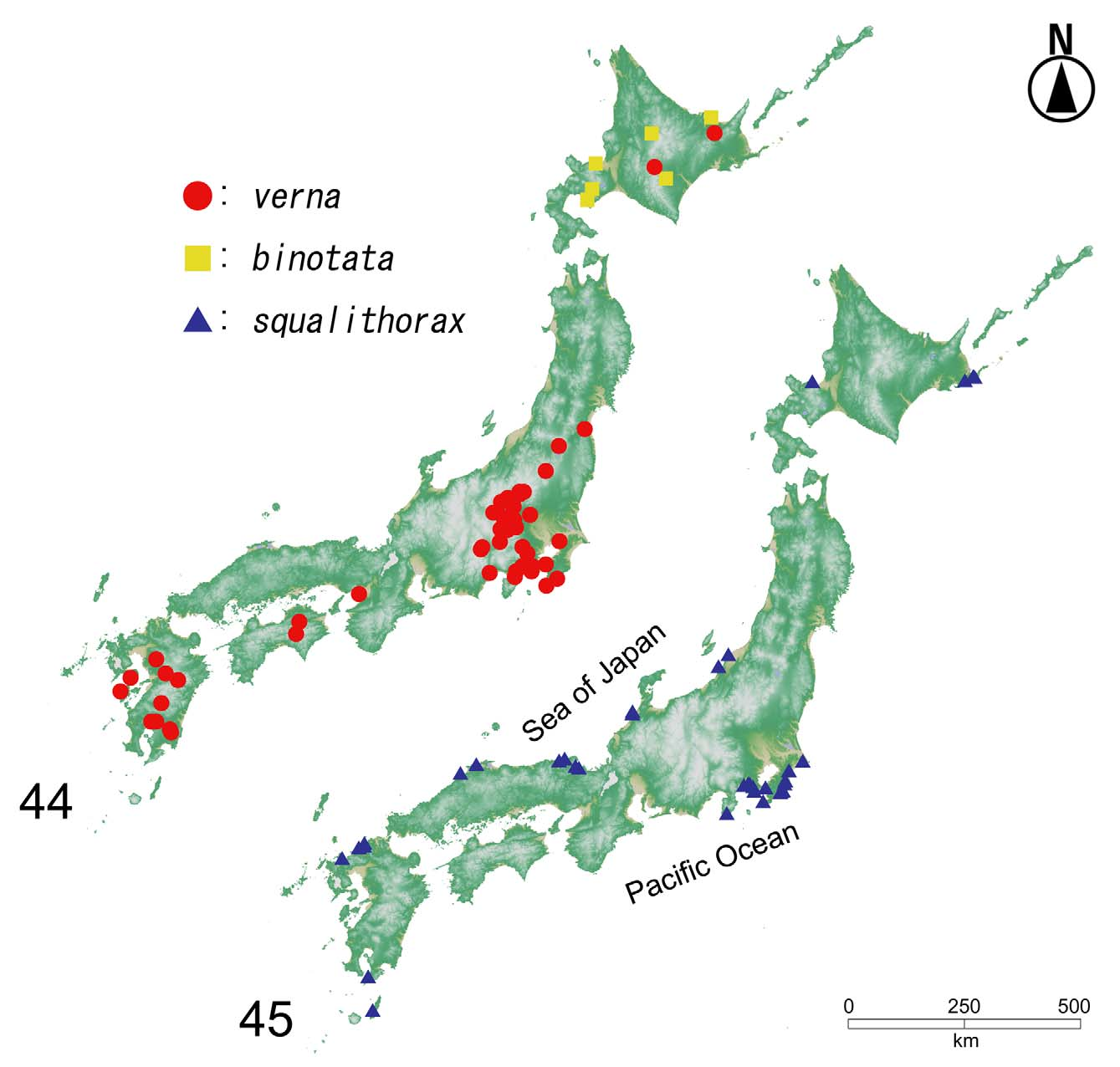
Confirmed distribution by the present study. [ JAPAN]: Hokkaidô, Honshû, Kyûshû, Tanega-shima.
Other localities in the literature. [ EAST ASIA]: South Korea (Ahn et al., 2000; Park & Ahn, 2004).
Remarks. Aleochara squalithorax is a rather common species in Japan but has a limited distribution globally. Until now, this taxon has only been recorded from Japan and South Korea (Smetana, 2004; Frank & Ahn, 2011). A recent study provided the first record of A. squalithorax from South Korea and briefly redescribed the species and provided a figure of the median lobe of the aedeagus of the male, based on Korean specimens (Ahn et al., 2000). The distribution of A. squalithorax in Japan is wide, from northeastern Hokkaidô (Nemuro) to southwestern Kyûshû (Tanega-shima). We provide new distributional records from mainland Kyûshû.
This species is considered a highly derived taxon of the subgenus Coprochara and it has attracted special attention from researchers (Bernhauer & Scheerpeltz, 1926; Assing, 1995; Maus, 1998). Thus, the line drawings of mouth parts, which are important characters in Aleocharinae that have never been described, are also provided in this paper.
Historical notes. This species was originally described as “ Aleochara squalithorax ” from “Hagi [Hagi-shi, western end of Honshû, Japan] (Fauvel), Hakodate [Hakodate-shi, Hokkaidô, Japan] (Lewis)” by Sharp (1888). Afterward, Fenyes (1920) included it within the peculiar subgenus Eucharina Casey, 1906 . A few years later, Bernhauer and Scheerpeltz (1926) transferred A. squalithorax to the newly established subgenus Skenochara Bernhauer & Scheerpeltz, 1926 as the type species, and it was the only species assigned to Skenochara . Assing (1995) raised this subgenus to generic rank (still only one species remained in the taxon). This paper was especially important because A. squalithorax was redescribed with figures and the male lectotype was designated. The systematic position of Skenochara was unclear, although Assing (1995) tentatively treated it as a distinct genus.
Maus and Ashe (1998) and Maus (1998) detected Skenochara with all apomorphies that characterize the subgenus Coprochara ; i.e., the existence of dorsal longitudinal rows on the pronotum, a strongly coiled spermatheca in the female, and a completely carinate mesoventrite. Therefore, Skenochara was synonymized under the subgenus Coprochara by Maus (1998). In addition to morphological evidence, molecular information supported this treatment (Maus et al., 2001; see Phylogeny of A. squalithorax ).
Bionomics. SY and MM collected the species at several different locations in Hokkaidô, Honshû and Kyûshû that faced both the Pacific Coast and the Sea of Japan ( Fig. 45 View FIGURES 44 – 45 ). All of the locations were sandy beaches, none of the areas were very small, and they generally had good environmental conditions ( Fig. 43 View FIGURES 42 – 43 ). Individuals were found from completely dry seaweed masses on beaches that were far from the shoreline from sites that were and close to coastal vegetation at Hamamatsu and Urayakotan in Hokkaidô; specimens were also collected from somewhat wet seaweed at Watari, Kyûshû ( Fig. 43 View FIGURES 42 – 43 ). Adults were observed and collected from early spring (March) to mid-fall (October), with the highest number of individuals being observed between April and May.
Host records. No information is available for A. squalithorax .
Phylogeny. Phylogenetic trees in Maus et al. (2001) show relationships among A. verna , A. binotata , and A. squalithorax . All three species were included in the Coprochara clade. Aleochara squalithorax formed a sister group with littoral A. (C.) sulcicollis Mannerheim, 1843 from North America. Microsculpture is present in Emplenota , Maseochara , Polystomota , and A. squalithorax , but none of the analyses supported a monophyletic clade of these taxa (Maus et al., 2001).
Color variation in elytra. Two patterns of elytral color variation were found. The gray type ( Fig. 3 View FIGURES 1 – 4 ) is apparently the typical form and the yellow type ( Figs. 4 View FIGURES 1 – 4 , 11 View FIGURES 5 – 13 ) is much rarer. We examined both types morphologically, but no differences were found between the two types. Thus, the authors concluded that the difference represents color variation within A. squalithorax . The yellow type accounted for only 12.6% of the total number of individuals collected (N=470). The ratio also showed considerable variation among locations. For example, no yellow-type individuals were found among 55 specimens at Sanriga-hama in Hokkaidô. In contrast, the yellow type accounted for 14.1% in Watari (data combined across small and large beaches), Kyûshû (N=156), and the highest ratio, 45.9%, was recorded in Katsuura, Honshû (N=37).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aleocharinae |
|
Genus |
|
|
SubGenus |
Coprochara |