
Myotis chiloensis ( Waterhouse, 1838 )
|
publication ID |
https://doi.org/ 10.1093/mspecies |
|
persistent identifier |
https://treatment.plazi.org/id/03D94C25-FFF7-9714-5EB4-FBE0FECEFD2A |
|
treatment provided by |
Carolina |
|
scientific name |
Myotis chiloensis ( Waterhouse, 1838 ) |
| status |
|
Myotis chiloensis ( Waterhouse, 1838) View in CoL
Chilean Myotis
Vespertilio Chiloensis View in CoL : Waterhouse, 1838:5. Type locality “islets on the Eastern side of Chiloe [Island],” Chile .
Vespertilio gayi Lataste, 1891:79 . Type locality “ Valdivia , Chile.”
Myotis chiloensis: Trouessart, 1904:94 View in CoL . First use of current name combination.
Myotis chiloensis dinellii: Miller and Allen, 1928:191 View in CoL . Type locality “ Tucuman, Catamarca Province, Argentina.”
Myotis chiloensis atacamensis Miller and Allen, 1928:192 View in CoL . Type locality “[San Pedro de] Atacama, Chile.”
Myotis chiloensis oxyotus Miller and Allen, 1928:193 View in CoL . Type locality “ Mount Chimborazo, Ecuador, at 9,000 to 10,000 feet altitude.”
Myotis chiloensis alter Miller and Allen, 1928:194 . Type locality “ Palmeiras , Parana, Brazil.”
Myotis chiloensis arescens Osgood, 1943:55 . Type locality “ Hacienda Limache , Province of Valparaiso, Chile.”
and M. c. alter as synonym of M. levis ( Barquez 2006) View in CoL . In South America, Cabrera (1958) assigned 5 subspecies to M. chiloensis View in CoL : M. c. chiloensis View in CoL , M. c. atacamensis View in CoL , M. c. alter, M. c. arescens , and M. c. oxyotus View in CoL . M. c. atacamensis View in CoL (currently M. atacamensis View in CoL — Barquez et al. 1999, Simmons 2005) and M.c. oxyotus View in CoL (currently M. oxyotus View in CoL — LaVal 1973) have both been elevated to specific status. M. arescens was synonymized under M. chiloensis View in CoL ( LaVal 1973; Gardner 2007) and M. c. alter was synonymized under M. levis ( LaVal 1973) View in CoL . The current classification of M. chiloensis View in CoL is based primarily upon morphological characters such as fur color and size ( Barquez et al. 1999; Gardner 2007). In Chile, no clear geographic
CONTEXT AND CONTENT. Order Chiroptera , suborder
Microchiroptera, family Vespertilionidae , subfamily Myotinae ,
genus Myotis , subgenus Leuconoe . Myotis chiloensis is cur-
rently treated as monotypic ( Simmons 2005; Wilson 2007). In the
past, 3 subspecies, distributed across central and southern Chile,
were assigned to M. chiloensis : M. c. chiloensis , M. c. arescens ,
and M. c. atacamensis ( Mann 1978) . In Argentina, 2 subspecies
were recognized by Cabrera (1958): M. c. alter and M. c. chi- Fig. 1. —An adult male Myotis chiloensis from Lampa, Metropolitan loensis, but LaVal (1973) considered M. chiloensis as monotypic region, Chile. Used with permission of the photographer Rodrigo del Piano. limit between M. chiloensis and M. atacamensis has been defined, and these species may be at least partially sympatric (Galaz and Yáñez 2006; Rodríguez-San Pedro et al. 2014). More research at the genetic level is necessary to define the limits between species and subspecies of Myotis in the southern cone of South America.
DIAGNOSIS
Geographically, Myotis chiloensis is sympatric with only 3 other Myotis species : M. atacamensis (Atacaman myotis) in Chile, and M. aelleni (southern myotis) and M. dinellii in Argentina. In Chile, M. chiloensis is distinguished from M. atacamensis because of the darker pelage of M. chiloensis ( Díaz et al. 2011) and the shorter forearm (mean) of M. atacamensis (31.3 mm versus 36.0 mm— Wilson 2007). The uropatagium of M. dinellii is pale in color and slightly hairy around the edge in contrast to M. chiloensis , which has a uniformly colored uropatagium with no hair extending to the edge. M. aelleni has tricolored fur on the dorsum and a longer forearm (37.7–41.2 mm) than M. chiloensis (37.0– 39.8 mm), which has bicolored fur on the dorsum, dark near the body and light at the tips ( Barquez et al. 1999; Díaz et al. 2011).
GENERAL CHARACTERISTICS
Myotis chiloensis (Fig. 1) is a small Vespertilionid bat endemic to the southern cone of South America. It shows latitudinal variation in fur color in relation to amount of solar radiation and precipitation ( Mann 1978; Galaz and Yáñez 2006). Northern populations are a pale ochraceous color, whereas populations of central Chile are pale brown, and southern populations show a coffee-brown fur (Galaz and Yáñez 2006).
Fur is bicolored and is about 4 mm in length (Barquez and Díaz 2009), dorsal fur is lighter and bicolored; individual hairs are pale brown or gray-brown, strongly contrasting with its lighter colored tips (Barquez and Díaz 2009). The short, soft fur on the dorsum of the uropatagium does not extend beyond the knees and the tail is entirely enclosed within the V-shaped uropatagium ( Díaz et al. 2011).
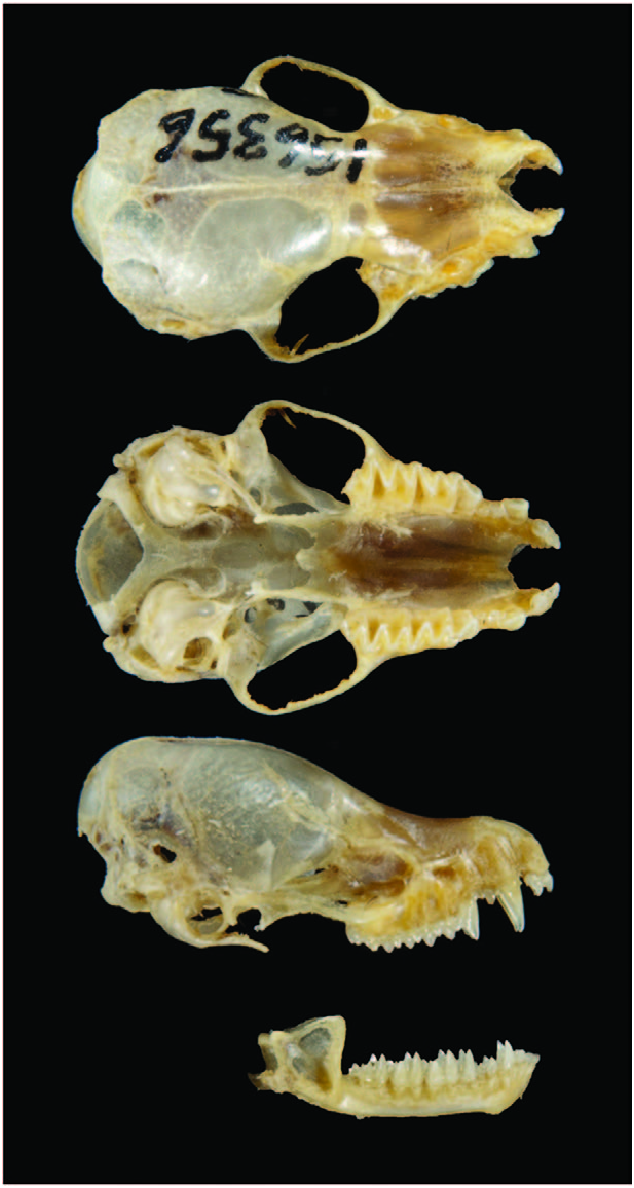
The skull of M. chiloensis is flat, with a ratio of greatest length of skull/braincase width equal to 2.0. Length of rostrum is almost as long as the braincase (Galaz and Yáñez 2006; Fig. 2 View Fig ). Mean measurements (mm ± SD) for 21 individuals from Chubut, Neuquén, and Rio Negro provinces in Argentina ( Barquez et al. 1999) were: condylobasal length, 13.6 ± 0.2; least interorbital breadth, 4.9 ± 0.1; zygomatic width, 9.1 ± 0.2; total length of skull, 14.4 ± 0.2; postorbital constriction, 3.8 ± 0.1; cranial box width, 7.1 ± 0.2; length of maxillary toothrow, 5.6 ± 0.1; palatal length, 7.1 ± 0.1; mastoidal breadth, 7.4 ± 0.3; length of mandibular toothrow, 5.8 ± 0.1; length of mandible, 10.8 ± 0.8; width across canines, 3.8 ± 0.0; and width across molars, 5.9 ± 0.1.
Mean (mm ± SD) morphometric measurements and body mass (g) for 9 individuals from Chile ( Ossa et al. 2010b) were: total length, 71.7 ± 6.2; length of tail, 28.7 ± 4.1; length of foot, 7.2 ± 0.9; length of ear, 9.5 ± 1.9; length of tragus, 4.8 ± 1.0; length of forearm, 38.5 ± 1.6; and body mass, 6.4 ± 0.7. Mean (mm ± SD) morphometric measurements and body mass (g) for 49 individuals from Chile ( Canals et al. 2001) were: body length, 47.7 ± 3.2; length of tail, 36.3 ± 6.0; length of forearm, 38.2 ± 1.4; and body mass, 6.7 ± 0.6. A mean body mass (g) for 17 males and 67 females from Chile was reported by Galaz and Yáñez (2006).
Mean measurements (mm ± SD) of 24 adult individuals from Chubut and Neuquén provinces, Argentina ( Barquez
1987) were: total length, 90.0 ± 4.28; length of tail, 37.4 ± 1.8;
length of foot, 8.4 ± 0.8; length of ear, 14.5 ± 1.2; length of forearm, 38.6 ± 0.8; and body mass, 7.5 ± 0.9 g. These values suggest that individuals in Argentina could be larger than Chilean individuals. Nevertheless, we recommend caution in interpreting these data because sample sizes are so small that the differences may be artifacts of the data rather than actual differences.
Recently, one of us (GO) measured 26 individuals captured in Lampa, Santiago de Chile. Mean measurements (mm ± SD) for 3 females were: total length, 81.4 ± 4.8; length of tail, 37.9 ± 3.3; length of foot, 7.4 ± 0.3; length of ear, 10.7 ± 0.3; width of ear, 5.8 ± 0.4; length of tragus, 5.0 ± 0.6; width of tragus, 1.7 ± 0.5; length of forearm, 39.1 ± 0.8; length of 5th finger, 47.6 ± 0.3; and body mass (g), 8.0 ± 1.0. Mean measurements for 23 males were: total length, 79.3 ± 3.2; length of tail, 38.9 ± 4.0; length of foot, 7.1 ± 0.9; length of ear, 11.5 ± 1.3; width of ear, 5.9 ± 0.7; length of tragus, 6.0 ± 0.9; width of tragus, 1.7 ± 0.2; length of forearm, 37.0 ± 1.6; length of 5th finger, 46.1 ± 1.3; and body mass: 6.2 ± 2.1.
DISTRIBUTION
Myotis chiloensis shares with Histiotus magellanicus
(southern big-eared brown bat), the southernmost known
distribution for any bat in the world ( Koopman 1967; Gardner
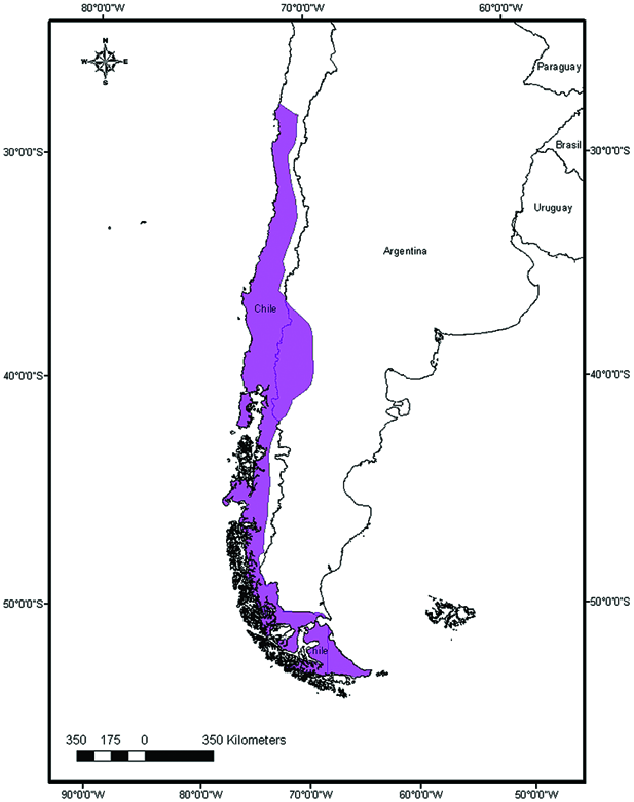
2007; Ossa and Díaz 2014). M. chiloensis is found only in Chile and Argentina (Redford and Eisenberg 1992; Simmons 2005; Wilson 2007; Fig. 3 View Fig ). In Chile, this species is described as occurring from Coquimbo (30°S — Galaz et al. 2009) to the northern shore of Navarino Island (55°S — Koopman 1967). The species probably occurs also inland on Navarino Island. In Argentina, M. chiloensis has been found in the provinces of Neuquén, Río Negro, Chubut, and Tierra del Fuego ( Koopman 1967; Mares et al. 1995; Barquez 2006; Barquez et al. 2012). Common habitats for this species are native forested areas, such as sclerophyllous forest in northern Chile and Temperate forest in their southern distribution ( Ossa et al. 2010b; Rodríguez-San Pedro and Simonetti 2013a). In Chile, no major geographical features limit the latitudinal distribution, but the gradual desertification to the north may limit its distribution where M. chiloensis is replaced by M. atacamensis ( Mann 1978) .
FOSSIL RECORD
Fossils of Myotis chiloensis have been recorded in 2 Holocene paleontological sites from Patagonia, Argentina ( Udrizar Sauthier et al. 2013).
FORM AND FUNCTION
Myotis chiloensis has a wing aspect ratio of 5.76 ± 0.16 95% CI (n = 37) and wing loading of 6.80 N /m 2 ± 0.23 95% CI (n = 47— Iriarte-Díaz et al. 2002 [mean ± SD]). Wing morphology of M. chiloensis is consistent with highly maneuverable flight needed to forage in cluttered habitats, including edge habitats and forest with clearings (Rodríguez-San Pedro and Simonetti 2013a). Its low aspect ratio and low wing loading indicate a high energetic cost and low flight speed, respectively ( Iriarte-Díaz et al. 2002). M. chiloensis is included in the guild of aerial insectivorous bats that forage in background-cluttered space (Schnitzler and Kalko 2001).
The mean basal metabolic rate of M. chiloensis is about 1.76 ± 0.28 ml O 2 g−1 h−1 ( Bozinovic et al. 1985). This value is within the range described for other insectivorous bats, but comparatively lower than those reported for frugivorous and nectarivorous ones ( McNab 1983). The small body size and low metabolic rate of M. chiloensis make it prone to fall into daily torpor ( Mann 1978; Bozinovic et al. 1985). M. chiloensis presents 2 responses to changes in environmental temperature: increasing its metabolic rate, maintaining an homeothermic temperature (36.6 ± 2.2°C), or decreasing its metabolic rate, maintaining the body temperature about 0.5°C over the environmental temperature ( Bozinovic et al. 1985). In summer, M. chiloensis has a single and short euthermic period of 2–3 h, followed by a long period of daily torpor or natural hypothermia ( Bozinovic et al. 1985), a pattern that apparenty is not shared by other congeneric species ( Pearson 1947; Kunz and Kurta 1988).
Dental formula is i 2/3, c 1/1, p 3/3, m 3/3, total 38, the highest recorded in bats ( Galaz et al. 2009). Upper incisors are well developed and similar in size, and there is a small diastema between canine and premolars ( Díaz et al. 2011).
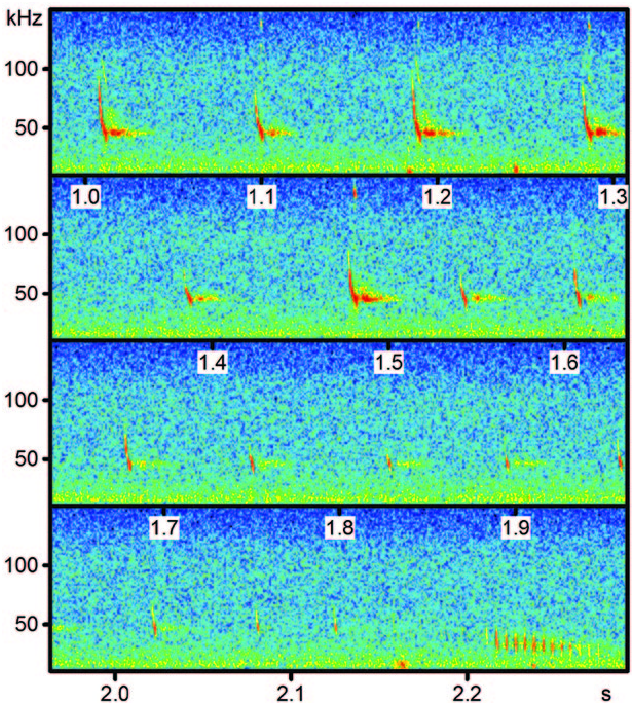
Echolocation calls of M. chiloensis contain a single harmonic and are characterized by a downward frequency modulation at the beginning of the signal followed by a narrowband quasi-constant frequency component ( Ossa et al. 2010a, 2010b; Rodríguez-San Pedro and Simonetti 2013b). Search calls consist of short (<4 ms) broadband signals sweeping down between 89 and 39 kHz with most energy at 47 kHz. Pulses are emitted at intervals of 95.06 ± 3.62 ms ( Ossa et al. 2010b; Rodríguez-San Pedro and Simonetti 2013b; Fig. 4 View Fig ).
ONTOGENY AND REPRODUCTION
Few data are available concerning pregnancy, lactation, or ontogeny. Myotis chiloensis , like other vespertilionids, is a monoestrus bat species (Pearson and Pearson 1989), and the age at 1st pregnancy is 10 months ( Galaz et al. 2009). As in other bats, the embryo develops in the right uterine horn ( Mann 1978). Females give birth to a single young per year at the beginning of summer ( Galaz et al. 2009). Mares et al. (1995) reported the capture of 1 pregnant female with a single fetus, which had a crown-rump length of 15 mm.
ECOLOGY
Myotis chiloensis is among the most common and widespread Chilean bats, occurring from the northern desert to the southern rainforest ( Mann 1978; Galaz and Yáñez 2006). This unimodal pattern clearly differs from the bimodal pattern exhibited by most insectivorous bats (Kunz and Fenton 2006). In arid and semiarid environments from northern Chile, M. chiloensis frequents watercourses for feeding ( Mann 1978). However, in south-central Chile, this species is associated with evergreen-deciduous forests and temperate forests, respectively ( Baker 1967; Galaz and Yáñez 2006; Rodríguez-San Pedro and Simonetti 2013a; Meynard et al. 2014). Exotic tree plantations (e.g., Pinus and Eucalyptus) are also used by M. chiloensis as foraging habitats in south-central Chile (Rodríguez-San Pedro and Simonetti 2013a; Meynard et al. 2014). In Argentina, specimens of this species were found beneath an abandoned bridge that crossed over a stream in riparian woodland within a Patagonian scrubland and roosting in an abandoned house in mixed stands of Nothofagus and Austrocedrus chilensis within the Valdivian Temperate Forest ( Mares et al. 1995; Gimenez et al. 2012).
A wide variety of roosting sites are used across Chile by M. chiloensis . In southern forests, for example, bats may use holes and fissures in tree bark as daily roosts, while artificial structures, caves, and abandoned mines provide ideal conditions for roosting in central and northern Chile ( Mann 1978). M. chiloensis roosts in a wide range of buildings in which they may use crevices in walls, attic spaces, or be found under tiles or other roofing materials ( Mann 1978; Galaz and Yáñez 2006; Ossa et al. 2010b; Giménez et al. 2012). Other smaller underground structures, such as rock crevices or ledges, may also be important for this species, particularly in the coastal area of the southern part of their range distribution (Galaz and Yáñez 2006).
BEHAVIOR
Myotis chiloensis feeds on insects, primarily nematocerans, that are captured in flight ( Galaz et al. 2009). Koopman (1967) reported this species feeding on Tipulidae flies in Southern Chile. Feeding activity of M. chiloensis begin at disk and last for about 3h, then bats return to roost ( Bozinovic et al. 1985; Galaz and Yáñez, 2006). This unimodal pattern clearly differs from the bimodal pattern exhibited by most insectivorous bats (Kunz and Fenton 2006).
In hibernacula, M. chiloensis is usually found with other bat species, especially the Brazilian free-tailed bat ( Tadarida brasiliensis ), the big eared brown bat ( Histiotus macrotus ), the small big eared brown bat ( H. montanus ), and the southern big eared brown bat ( H. magellanicus — Mann 1978; Mares et al. 1995).
GENETICS
The diploid number (2n) for Myotis chiloensis is 44 chromosomes, and the number of arms of the autosomal complement (fundamental number, FN) is 50. The X chromosome is submetacentric (Galaz and Yáñez 2006).
CONSERVATION
Myotis chiloensis is regarded as “Least Concern” by the International Union for Conservation of Nature and Natural Resources Red List of Threatened Species because it is a common species; however, its population trend is unknown (Barquez and Díaz 2008). There are no major threats to this species throughout its range.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myotis chiloensis ( Waterhouse, 1838 )
| Ossa, Gonzalo & Pedro, Annia RODRíGUEZ-San 2015 |
Myotis chiloensis arescens
| OSGOOD, W 1943: 55 |
Myotis chiloensis: Trouessart, 1904:94
| TROUESSART, E 1904: 94 |
Vespertilio gayi
| LATASTE, F 1891: 79 |
Vespertilio
| WATERHOUSE, G 1838: 5 |