
Hydrodendron chilense Galea & Schories
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3852.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:7DE3BCBA-E5F0-4F0D-B2FD-B5B59E4DAE51 |
|
DOI |
https://doi.org/10.5281/zenodo.6143187 |
|
persistent identifier |
https://treatment.plazi.org/id/03D92A2C-4770-FFC5-FF51-868358E0A1C1 |
|
treatment provided by |
Plazi |
|
scientific name |
Hydrodendron chilense Galea & Schories |
| status |
sp. nov. |
Hydrodendron chilense Galea & Schories , sp. nov.
( Fig. 5A–E View FIGURE 5. A ‒ E , Table 5 View TABLE 5 )
Material examined. Corral, Chaihuin/Huiro, lat. -39.95000, long. -73.61667, 17.x.2011, 8‒ 12 m, sample 36: a male colony on seaweed (holotype: MHNG-INVE-86227). France, Port d’Alon creek, lat. 43.146369, long. 5.707545, 20.v.2009, 0.5 m, sample HRG-0020: a female colony of H. mirabile ( Hincks, 1866) on seaweed. France, Cap Taillat, lat. 43.174040, long. 6.640635, 09.vi.2011, 0.5 m, sample HRG-0567: a male colony of H. mirabile on seaweed. France, Martinique, Anse Dufour, lat. 14.527162, long. -61.090033, 12.i.2014, 0.5 m, sample M277: several colonies, some fertile, of H. mirabile on seaweed.
Description. Colony arising from a creeping, branching, anastomosing hydrorhiza; perisarc with irregular, thick, internal projections. Hydrocauli upright but small, not surpassing ca. 3 mm in height. Stems monosiphonic, divided into internodes by transverse constrictions of the perisarc. Internodes broadly cylindrical, of varied length, each with a proximal bulge, with or without a lateral apophysis supporting the following internode or a hydrophore, and with or without a nematotheca; perisarc moderately thick, surface smooth to locally wrinkled. Branching pattern clearly sympodial, with additional internodes or hydrophores given off at acute angles from the apophyses of preceding internodes; apophyses not in one plane, but arising from any side of the stems. Hydrophores of varied length, with a basal bulge, widening gradually distally to merge into hydrotheca; both separated by a thin diaphragm projecting into the lumen of hydrophore. Hydrotheca tronconical, obviously deep, rim flaring, margin conspicuously everted. Ring of desmocytes irregular, placed at a fair distance above diaphragm. Hydranths reduced to mere buds, possibly due to reproductive exhaustion. Nematothecae irregular and scarce, given off from both the stolon and some stem internodes or hydrophores; beaker shaped, rather large and deep, with a distinctly everted rim; nematophores long (up to ca. 2 mm) and slender, with solid core and a large, distal capitulum. Gonothecae, male in the present material, borne abundantly on the stolon; elongated tubular for most of their length, tapering abruptly proximally into a short, indistinct pedicel; truncate distally above a narrowed neck; walls provided with 7–8 distant, spiral grooves.
Remarks. The new species is characterized, among others, by the presence of spirally twisted gonothecae. Four congeners, namely H. alternatum ( Fraser, 1938) , H. expansum ( Fraser, 1948) , H. mirabile ( Hincks, 1866) , and H. sympodiformis Millard & Bouillon, 1974 , share the same character. Both H. alternatum and H. expansum were described from the eastern Pacific. Similarly to H. chilense , the internodes of the former are given off sympodially at sharp angles from the preceding internodes, but there is reportedly a characteristic alternate arrangement of the hydrothecae along both sides of the stems and, when present, side branches ( Fraser 1938), a situation not met in the new species. In addition, H. alternatum is further distinguished through the apparently exclusive and abundant occurrence of nematothecae on its stems [“commonly one to each pedicel” (sic)], and its uncommon ability to give rise to gonothecae, not only from the stolon, but also from the hydrophores and the stem internodes.
Unlike H. chilense , H. expansum is a species with polysiphonic stems, collinear internodes delimited by oblique nodes, and large gonothecae with strongly ringed walls, which are given off “from the fascicled portion of the stem, projecting at right angles to it” ( Fraser 1948).
Hydrodendron chilense shares several features with H. mirabile , among which the sympodial growth pattern of the stems15. The material in hands was compared with three samples that we assign to H. mirabile : a male and a female16 colony from the Mediterranean ( Fig. 5 View FIGURE 5. A ‒ E F–J), as well as a male colony from Martinique ( Fig. 5 View FIGURE 5. A ‒ E L– O). Although all exhibit a sympodial growth, in H. chilense the hydrothecae are much deeper and have a strongly everted rim, the nematothecae are bigger and their rim is equally everted, and the gonothecae are more elongated and reach as much as two times the length of their Caribbean and Mediterranean counterparts (see measurements in Table 5 View TABLE 5 ). In all available accounts dealing with fertile material [notably Rho & Park (1980); Hirohito (1974, 1995)] attributable, with little doubt, to H. mirabile , the gonothecae never reach the huge size seen in the Chilean species. The Vema material studied by Millard (1966b), however, possesses gonothecae whose size is comparable to that observed in H. chilense , but their walls are only “lightly annulated”. In addition, their trophosome is said “similar to the South African material” that she described earlier as having “the hydrophores alternately to right and left” ( Millard 1957), thus differing from the typical growth pattern of H. mirabile .
Hydrodendron sympodiformis is almost indistinguishable morphologically from the specimens of H. mirabile examined by us [compare Millard & Bouillon’s (1974) Fig. 5 View FIGURE 5. A ‒ E with our Fig. 5 View FIGURE 5. A ‒ E F, I, L, O], and its measurements fit well those of the Mediterranean and Martinican samples (see Table 5 View TABLE 5 ). For this reason, we suspect that it could be a junior synonym of Hincks’ species.
The cnidome composition of all specimens studied here, as well as that of a sterile congener from Guadeloupe (see Galea 2010a, p. 13, Fig. 4A–D View FIGURE 4. A ‒ F, M ), comprises six capsules: a small ( Fig. 5E View FIGURE 5. A ‒ E 3, K3, P3, Q3) and a very large ( Fig. 5E View FIGURE 5. A ‒ E 1 View FIGURE 1. A, B , K1, P1, Q1) pseudostenotele, as well as four microbasic mastigophores ( Fig. 5E View FIGURE 5. A ‒ E 2 View FIGURE 2. A, B , 4 View FIGURE 4. A ‒ F, M –6, K2, 4–6, P2, 4–6, Q2, 4–6). The shape and size of the capsules appears uniform in all specimens with sympodial colonies (compare Fig. 5E View FIGURE 5. A ‒ E , K & P), while a difference in the size of the large pseudostenoteles ( Fig. 5 View FIGURE 5. A ‒ E Q) could be noted in the species from Guadeloupe, whose structure of stems is radically different, suggesting that, in some cases, species-specific differences may occur.
It has been shown earlier (Galea et al. 2012) that the types of capsules within the genus Nemalecium Bouillon, 1986 View in CoL do not change, but their sizes vary to a certain extent among species. Conversely, in Halecium Oken, 1815 View in CoL there is a significant variation in the shape, type and size of the capsules, as illustrated with the species discussed above.
Etymology. The new species is named after the country of its origin.
Distribution in Chile. Only known from Corral.
15. The stems of H. mirabile were originally described as “rudely annulated at the base, and jointed at intervals throughout”, with the “branching […] usually consisting of a single division of the stem”, at most two ( Hincks 1866). A modern illustration of the neotype (BMNH 1899.5.1.169) is given by Cornelius (1995a, Fig. 73C), who showed the characteristic sympodial growth pattern of the colonies. The description and illustration of the gonotheca goes back to Hincks (1868). It is worth underlining that the description of H. mirabile provided by Cornelius (1975) resulted undoubtedly from pooled data based, among others, on material dealt with earlier by Ritchie (1908, as Ophiodissa caciniformis ) and Millard (1957, 1966b). In Cornelius’ (1975, 1995a) view, widely held by subsequent authors, H. caciniformis represents not more than a junior synonym of H. mirabile , and it appears that he mainly focused his attention to the similarities between them, rather than to certain, though not less important differences that are related to their respective growth patterns. Indeed, his illustration ( Cornelius 1975, Fig. 14A) of the paralectotype of O. caciniformis shows a species with almost collinear internodes delimited by slightly oblique nodes, thus differing from the sympodial growth pattern characteristic of Hincks’ species. In addition, the type material of H. caciniformis is sterile. A careful check of the relevant literature shows that specimens with both sympodial [e.g. Babić (1913), García Corrales et al. (1978), Rho & Park (1980), Hirohito (1974, 1995), Ramil Blanco & Iglesias Diaz (1988), Altuna (1994), Peña Cantero & García Carrascosa (2002)] and alternate [e.g. Millard (1957, 1966b), Ralph (1958), Vervoort (1959), Gili (1986), Medel et al. (1998), Vervoort & Watson (2003)] growth patterns were equally included in either H. mirabile or H. caciniformis . It therefore becomes obvious that some of the records cited above, especially certain based on sterile material, are either incorrect or belong to unidentifiable species.
- total length 1350–1720 (♂) 830–855 (♂) 670–830 (♂) 630–800 (♀) – ca. 1120 (♀)
FIGURE 6. A, B: Sertularella asymmetra sp. nov. —colony fragment (A); hydrotheca in lateral (B1) and apical (B2) views. C: Sertularella blanconae El Beshbeeshy, 2011 —colony fragment. D, E: Sertularella curta sp. nov. —colony fragment (D); hydrotheca in lateral (E1) and apical (E2) views. F, H: Sertularella pauciramosa sp. nov. —colony fragment (F), and comparison with S. polyzonias (Linnaeus, 1758) (G); four hydrothecae (H). Scale bars: 200 µm (B, E, H), 1 mm (A, C, D, F, G).
FIGURE 7. A: Sertularella asymmetra sp. nov. —female (A1) and male (A2) gonotheca; B: Sertularella blanconae El Beshbeeshy, 2011 —male gonotheca. C: Sertularella curta sp. nov. —male gonotheca. D: Sertularella pauciramosa sp. nov. —male (D1) and female (D2) gonotheca. E, F: Symplectoscyphus flexilis Hartlaub, 1901 —colony fragment (E1) and hydrotheca (E2); gonotheca (F). G‒J: Symplectoscyphus semper sp. nov. —stem fragment with beginning of side branch (G1), and portion of cladium (G2); hydrotheca (H); gonotheca (I), and details of the distal part (J). K‒M: Symplectoscyphus cf. subarticulatus ( Coughtrey, 1875) —colony fragment (K); hydrotheca (L); gonotheca (M). Scale bars: 200 µm (H, L), 500 µm (A‒D, F, I, J, M), 1 mm (E, G, K).



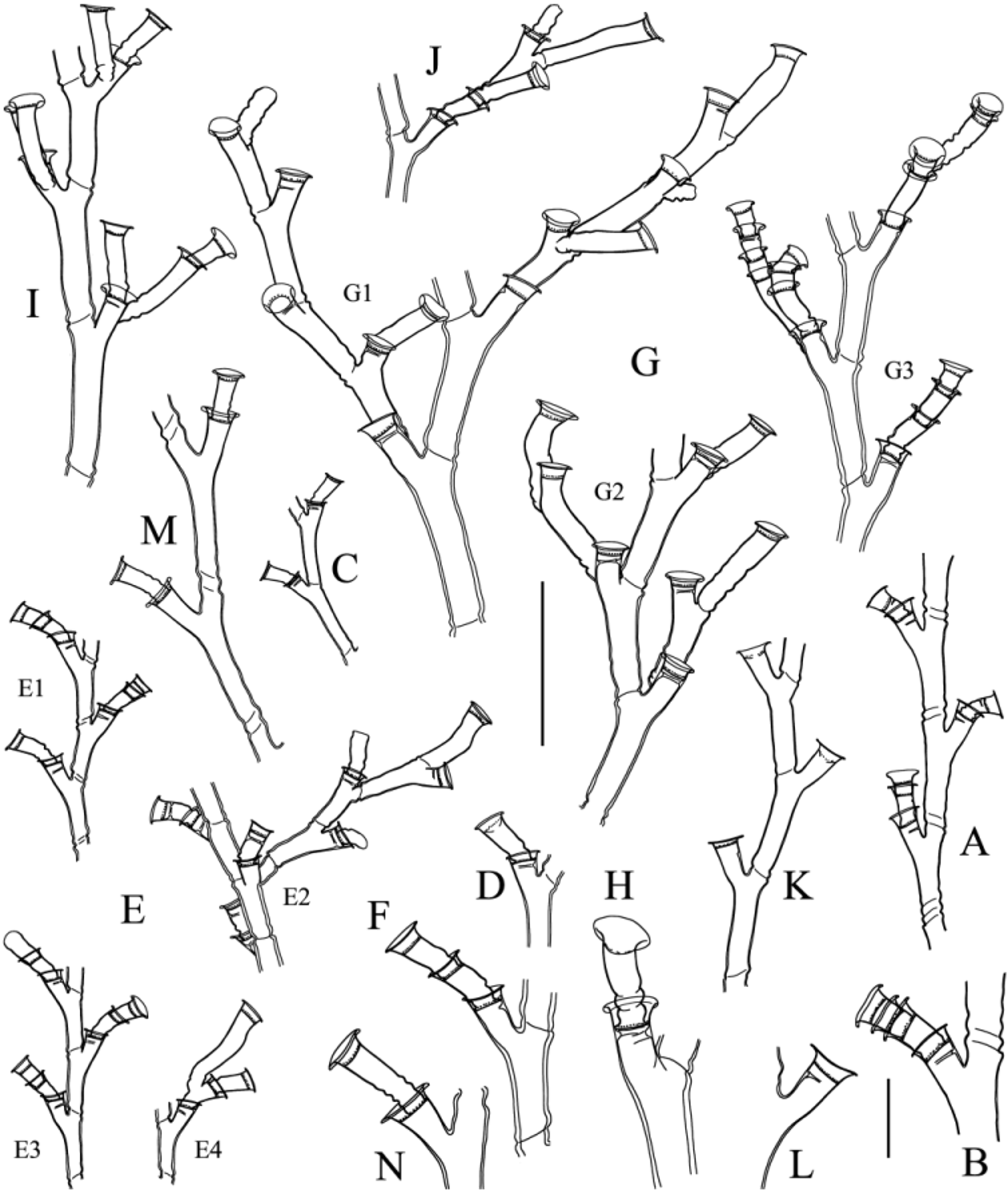
TABLE 5. Morphometric comparison of Hydrodendron species (dimensions in µm).
| Species | H. chilense | H. mirabile | H. mirabile | H. sympodiformis Hydrodendron sp. |
|---|---|---|---|---|
| Reference(s) | Present study | Present study | Present study | Millard & Bouillon Galea (2010a), (1974) present study |
| Distribution | Chile | Mediterranean | Martinique | Mozambique Guadeloupe |
| Internodes | ||||
| - length | 100–315 | 120–405 | 115–270 | – – |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hydrodendron chilense Galea & Schories
| Galea, Horia R., Schories, Dirk, Försterra, Günter & Häussermann, Verena 2014 |
Nemalecium
| Bouillon 1986 |
Halecium
| Oken 1815 |