
Seira dowlingi
|
publication ID |
https://doi.org/ 10.5281/zenodo.180444 |
|
DOI |
https://doi.org/10.5281/zenodo.6229070 |
|
persistent identifier |
https://treatment.plazi.org/id/03D8A82D-FFF9-FFCA-FF3D-F9FDFDEAFB9A |
|
treatment provided by |
Plazi |
|
scientific name |
Seira dowlingi |
| status |
|
No scales are present on first instar larvae.
Scale distribution on antennae. Two scales appear on the dorsal base of Ant. I–II during 2nd Inst.; there are no scales on Ant. III–IV or on the ventral side of any segment. The distribution of scales remains unchanged during 3rd Inst. and expands minimally at 4th Inst. By 5th Inst. scales cover all the dorsal surface of Ant. I, the basal half of Ant. II, and the basal fifth of Ant. III. In adults, two thirds of the dorsal surface and somewhat less than half of the ventral surface of Ant. II are covered with scales; Ant. III has scales dorsally on the basal fourth and ventrally on the basal fifth; scales are absent from Ant. IV and the ventral surface of Ant. I.
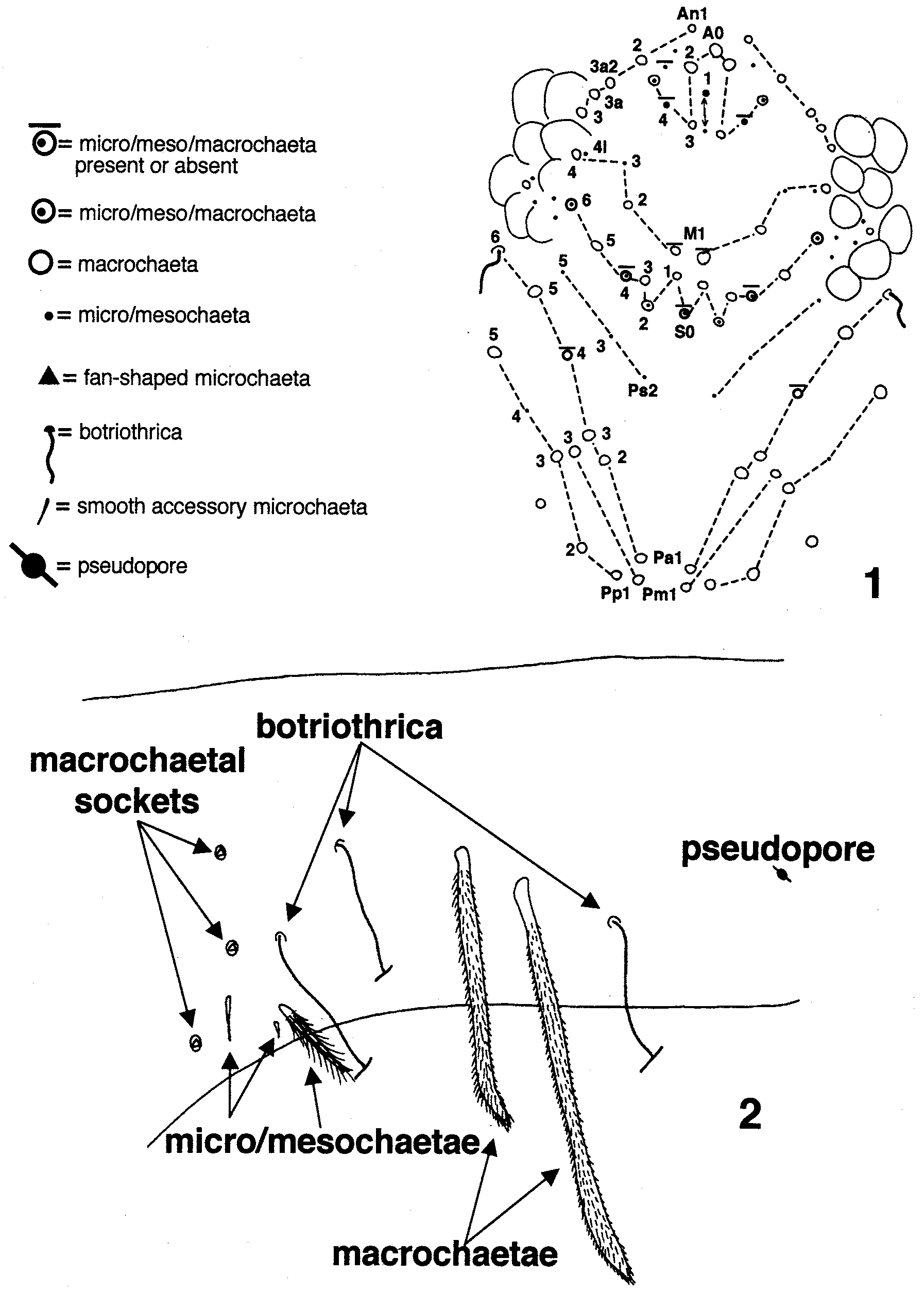
Ant. I botriothrica and sensilla. The first instar has three ciliate setae and is devoid of botriothrica (Fig. 13). The number of setae and sensilla increases during 2nd -3rd Inst. (Figs 14, 15), but no botriothrica appear until 4th Inst. (Fig. 16). A second botriothricum is added during 5th Inst. (Fig. 18). Adults appear to be dimorphic in the number of botriothrica: males have four, females three (Figs 19, 20).
Ant. II botriothrica and sensilla. The first instar (Fig. 9) has one botriothricum and some ciliate setae, but no other sensilla. At 2nd Inst. (Fig. 10) the number of ciliate setae increases, and a thin-walled, tapered sensillum is added. A second botriothricum appears at 3rd Inst. (Fig. 11). The third botriothricum is added at 4th Inst. (Fig. 12). In subsequent instars the relative distance between the three botriothrica changes, but only ciliate setae and sensilla are added. Short, conic setae associated with the botriothrica appear by 5th Inst. (arrows in Fig. 17), increasing numerically during subsequent instars (Fig. 21).
Ocular setae. Most adults have five setae in the eyepatch, probably homologous to setae p, q, r, v, and t in Lepidocyrtus ( Mari Mutt 1986a). Seta s is asymmetrically present in one specimen ( Fig. 27 View FIGURES 27 – 30 ). Setae p, r, and t are primary (Fig. 24). Additional setae first appear during 4th Inst., but are unstable: of seven specimens examined four have only the primary setae, one specimen adds v, but two others add q instead. All setae are present in all specimens beginning at 5th Inst.
Labium. In the labial triangle of 1st and 2nd Inst. (Fig. 23) the anterior setae are smooth, the posterior setae are ciliate, and seta r is absent (M1oEA1–2). Only the presence or absence of setae M2 and r varies during postembryonic development. In 3rd Inst., one specimen has both setae; five specimens in 4th Inst. have M2 r, one has only r (short, smooth and conic) and one lacks both. The final chaetotaxy is reached at 5th Inst., all anterior setae are smooth macrochaetae and the posterior setae, except r, are ciliate macrochaetae (M1 M2 r E L1–2 A1–2). All adults have the complete chaetotaxy.
Scale distribution on legs. The pro- and mesothoracic legs lack scales during 2nd Inst., but there is a single scale, distally on the external side of the metafemur and metatibiotarsus. Scales appear on the meso- and metacoxae in 3rd Inst. By 4th Inst. there are scales on all segments of all legs, but on the first two pairs the scales do not cover all surface. During 5th Inst. all legs have a similar density of scales.
Scale distribution on collophore. Scales (2–3) first appear laterally on the anterior face during 4th Inst. By 5th Inst. both faces of the collophore have scales, concentrated on the central region of the organ, but they are more dense on the anterior face. The density of scales increases on both faces in adults (with respect to 5th Inst.), but the density remains higher on the anterior face.
Scale distribution on manubrium. The dorsal manubrial face lacks scales in 2nd Inst., but on the ventral face there is one line of scales along the lateral margins. The density of ventral scales increases with every instar, creating a uniform cover, but dorsal scales do not appear until 5th Inst. Adults have one line of scales along the margins of the dorsal face.
Scale distribution on dens. During 2nd Inst. scales cover two thirds of the ventral dental face, and by 3rd Inst. they reach the base of the uncrenulate area anterior to the mucro. No scales are present on the dorsal dental face of adults.
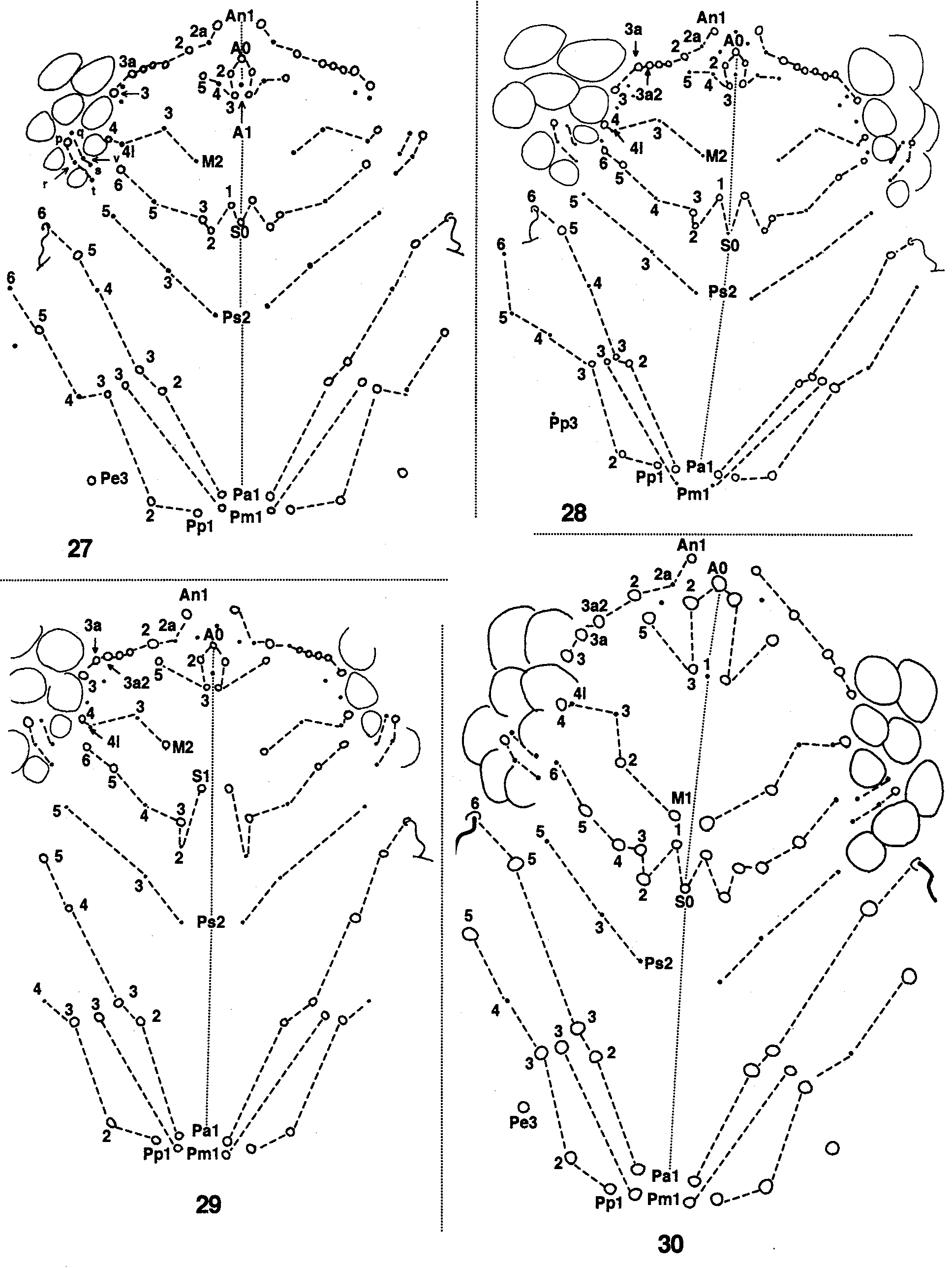
Dorsal head chaetotaxy. The five rows of anterior setae (An, A, M, S, and Ps, Fig. 1 View FIGURES 1 – 2. 1 ) are already present in 1st Inst. (Fig. 24). Rows S and Ps have the full complement of setae already during 1st Inst. Post-embryonic changes on these rows are limited to the displacement of setae with respect to each other or their transformation into macrochaetae. The macrochaetotaxy of the posterior region is weakly developed in S. dowlingi ; adults have four rows of setae and all are present in 1st Inst.
Row An. Adults have six to nine setae on this row ( Fig. 27 View FIGURES 27 – 30 ) but only An1, 2, 3 are primary (Fig. 24). An1–3 are microchaetae during 1st Inst., but are transformed into acuminate macrochaetae by 2nd Inst.. A fourth macrochaeta (An3a) appears between An2-3 at 2nd Inst. (Fig. 25). Microchaeta An2a and a macrochaeta anterior to An3a appear by 3rd Inst. (Fig. 26), but they are not stable; the seta anterior to An3a is missing in four specimens of seven studied, whereas An2a appears as a microchaeta or a scale. Both setae are fixed at 4th Inst. A smooth microchaeta posterior to An3 appears at 4th Inst., but it is not fixed, remaining variable in the adults ( Fig. 27 View FIGURES 27 – 30 ). All additions of macrochaetae to this row subsequent to 5th Inst. occur between An2 and An3a and seem to be added from posterior to anterior. The number of macrochaetae in this region is variable in the material studied, varying from one to three at 5th Inst. and three to four in adults. The microchaeta antero-internal to An3 appears after 5th Inst.
Row A. Adults have four paired (A2–5) and two unpaired (A0–1) setae in this row. Setae A0–3 and A5 are primary. Secondary seta A4 (Fig. 25) appears at 2nd Inst.
Row M. Adults of S. dowlingi have setae M2, M3, M 4i, and M4 ( Fig. 27 View FIGURES 27 – 30 ). During 1st Inst., M0, M2, M3, and M4 are microchaetae (Fig. 24), but M1 is absent. At 2nd Inst. seta M0 disappears (is not present even as a scale) and M 4i appears (Fig. 25). M4 becomes an acuminate macrochaeta at 3rd Inst. (Fig. 26). There are no changes in this row after 3rd Inst.
Row S. In adults of S. dowlingi this row includes nine macro- and two microchaetae arranged as 5+5 setae plus the unpaired S0 ( Fig. 27 View FIGURES 27 – 30 ). At 1st Inst. macrochaetae S0–3 are arranged in two straight rows, with S3 horizontally aligned to S1. By 2nd Inst. (Fig. 25), S3 has moved posteriorly, closer to S2 than S1. S5 remains a microchaetae in the adult. S6 first appears as a macrochaeta in some specimens during 3rd Inst. (Fig. 26) and is fixed by 4th Inst. S4 is absent in all instars.
FIGURES 9–20. Seira dowlingi ; 9–12, Antennomere II; 9, 1st instar; 10, 2nd instar; 11, 3rd instar; 12, 4th instar; 13–16, Antennomere I; 13, 1st instar; 14, 2nd instar; 15, 3rd instar; 16, 4th instar; 17, 5th instar antennomere 2, arrows identify conic setae; 18–20, Antennomere I; 18, 5th instar; 19. Adult male; 20, Adult female.
FIGURES 21–26. 21, Seira dowlingi , adult antennomere 2; 22, S. blanca , adult antennomere 2; 23–26, S. dowlingi ; 23, Labial triangle, 1st instar; 24–26. Dorsal head chaetotaxy; 24, 1st instar; 25, 2nd instar; 26, 3rd instar.
Row Ps. Adults have 3+3 microchaetae, all primary. The only change during postembryonic development is a widening of the gap between Ps3 and Ps5 (cf. Figs. 24, 27).
Row Pa. Adults of S. dowlingi have one botriothricum, four macro- and one microchaetae; the botriothricum and four macrochaetae are primary (Fig. 24). The final chaetotaxy is completed at 2nd Inst. (Fig. 25) with the appearance of microchaeta Pa4. Macrochaeta P0 of Mari Mutt (1986b) corresponds to Pa 5 in the present nomenclature system.
Row Pm. Adults ( Fig. 27 View FIGURES 27 – 30 ) retain the number of setae found in 1st Inst.
Row Pp. The organization found in adults is reached at 2nd Inst. with the addition of microchaeta Pp6 and macrochaeta Pp7 (Fig. 26). One specimen at 2nd Inst. has two (transient?) macrochaetae of unclear homology between Pp4–5 (Fig. 25). Macrochaeta P2 of Mari Mutt (1986b) corresponds to Pp 5 in the present nomenclature.
Mesothorax. 1st Inst. lacks setae m3 and p7 (Fig. 31); a7, m5, m7, p4, and p6 are ciliate micro- or mesochaetae, all others are ciliate macrochaeta. Nineteen secondary setae are added by 2nd Inst. (Fig. 32), most of them as ciliate macrochaeta, none of the seven primary microchaetae changes into a macrochaeta. There is much variation in the shape of setae in the posterior row, in area Pm. The most common arrangement in this area includes three groups (see Fig. 36 for group composition): group A has two macrochaetae (2M), group B has two macro- and one microchaetae (2M, 1µ), and group C has three macrochaetae (3M). Other arrangements observed among the ten individuals studied are (3M) (1µ) (3M); (2M) (1M, 1µ) (3M); (2M) (2M, 1µ) (4M).
Instar 3 (Fig. 33) shows a general increase in the number of macrochaeta in the collar but only seven new setae appear posterior to the collar: macrochaetae a5p, a 5i and p 1i; and microchaetae a2p, m 4i, m4ip and p1ip2. The pattern in area Pm at 3rd Inst. is (1µ, 3M) (3M) (3M).
During 4th Inst. the collar retains the multiplet structure, but the homology of setae is difficult to discern, especially in the lateral area. Most specimens do not show the clear organization shown in Fig. 34. Five new setae are added, or change from µ to M, posterior to the collar (Fig. 34): macrochaetae a 5i 2, p1ip, p2ea, p3p, and microchaeta m 1i 2. The formula of area Pm is (1µ, 4M) (3M) (5M); variation was limited to PmC (4M, 1µ). The group of m1–2 is unstable at this instar, m1a2 is missing in three specimens, and m1a is a macrochaeta in two specimens. Other changes at this instar are the anterior displacement of m4p (cf. Figs 33–34) and the transformation of m 4i into a macrochaeta. In 5th Inst. the chaetotaxy of the area posterior to the collar is identical to that in adults, but the collar is only moderately more densely packed with macrochaetae than during 4th Inst. (Fig. 35). The number of collar setae continues to increase in successive molts and the largest adults have considerably more setae than specimens in 5th Inst.; the multiplet system is obliterated by the large density of macrochaetae in this and subsequent instars. Adults have (1µ, 5M) (3M) (5M) setae in region Pm (Fig. 36): four macrochaetae in groups m1–2 and a5, and three in group m4. The base of m1 is posterior to m2; m 4i is often posterior to m4p; and p3, p3p and p2e form a densely packed diagonal.
Setae identified here as a 5i and a 5i2 may be homologous to those included in generatrix a4 and a 4i in other entomobryids ( Szeptycki 1979), but their homology is unclear. Seta a 5i may correspond to a4p2, and a 5i 2 to a4ip (cf. Fig. 34 and Figs 80–81 View FIGURES 80 – 83 in: Szeptycki 1979), but by 5th Inst. both setae are displaced towards the segmental midline and a 5i is aligned with the multiplet a 4i. In adults these setae are very distant from the collar. Since additional setae appear between a 5i and a 5i 2 and their presumptive generatix, it is impossible to establish a reasonable homology with apparently corresponding setae in adults of other genera.
Metathorax. The only primary setae absent at 1st Inst. are m2, m3 and p7 (Fig. 41). Seta m1 and the supplementary seta external to a7 (asterisk in Fig. 41) are smooth, other setae are ciliate. One of the seven individuals examined carried a supplementary seta of uncertain homology external to a3 (arrow in Fig. 41). Seta m6p and macrochaeta p2a appear at 2nd Inst. (Fig. 42). At this instar, m1 moves to the inside of a line drawn between a1 and p1; a4 and a5 move posteriorly, just above the pseudopore and almost in line with p3; a6 also FIGURES 31–36. Seira dowlingi , mesotergal chaetotaxy; 31, 1st instar; 32, 2nd instar; 33, 3rd instar; 34, 4th instar; 35, Collar of 5th instar; 36, Adult, excluding collar.
migrates posteriorly, aligning itself with p5 and p6. The number of setae present in the adult is reached at 3rd Inst. (Fig. 43), with the addition of microchaeta a1a. After 3rd Inst., chaetotacic changes are restricted to microchaetae length reduction and their progressive distancing from the macrochaetae (cf. Figs 41, 44). Adults have one microsensillum external to m7, nine microchaetae, two acuminate mesochaetae (dots corresponding to m7 and m6p, Fig. 44), and ten truncate macrochaetae. An interesting aspect of the development of this segment is that all macrochaetae on the anterior row external to a2 move posteriorly into alignment with macrochaetae on the posterior row. The most dramatic case is the migration of a6, which in the adult is located posterior to p6.
Abdomen I. Setae absent at 1st Inst. are a7, m1, m7, and all setae on the posterior row, except p5 and p6 (Fig. 51). Only m2–4 are truncate macrochaetae, although the sockets of a1, m6 and p6 are large and could be confused with those of macrochaetae. All setae are ciliate, except the setula external to a6. At 2nd Inst. a new macrochaeta appears between m4 and m3 (Fig. 52). This seta appears almost directly posterior to a3, suggesting that it is homologous to m 4i present in some species of scaleless Orchesellinae , Entomobrya , Homidia and Himalanura ( Szeptycki 1979) . There are no changes in the structure of setae with respect to 1st Inst., but a5 and m5 are more posterior and form a row with p5 and p6 (respectively), and as in other entomobryids, a1 moves posterior to the pseudopore. The final set of setae present in the adult is completed at 3rd Inst. with the addition of a1a (Fig. 53). At 4th Inst. all microchaetae are smooth. In adults (Fig. 54) setae a1a, a2, and a3 do not reach the sockets of the macrochaetae, and a6 and p6 are aligned, forming a column with the lateral setula.
Abdomen II. The primary setae absent at 1st Inst. are a4, m1, and p1–3 (Fig. 60). All setae are ciliate, except as and se, and a2 is inserted anterior to the pseudopore. At 2nd Inst. macrochaeta m3e and microchaeta m3ea are added, whereas a1, m4, and p4 are transformed into scales (Fig. 61). Seta m7 moves anteriorly forming a row with a7. No primary setae are lost after 2nd Inst. Some of the fan-shaped setae around the botriothrica appear at 3rd Inst. (triangles in Fig. 62), but their number varies from 2–4 around m2 and 1–3 around a 5. I consider the fan-shaped setae posterior to m2 homologous to a2p in Pseudosinella , although it appears at 3rd Inst. in S. dowlingi and at 2nd Inst. in P. alba ( Packard, 1873) . Based on their relative position, the fanshaped setae anterior to m2 and posterior to a 5 may be re-diferentiated setae a1 and m4, respectivly. All fan setae of the botriothrical complexes present in the adult appear by 4th Inst. (Fig. 63). By 4th Inst. seta a2 is consistently inserted posterior to the pseudopore. At 5th Inst. seta a2 is further displaced posteriorly. In adults all microchaetae, except p6, are considerably shorter than at 1st Inst. (Fig. 64).
Abdomen III. The primary setae absent at 1st Inst. (Fig. 69) are a4, m1, p1, and p2. Setae as, d2, and se are smooth; a1, m3, pm6 and p6 are ciliate macrochaetae; other setae are ciliate. Changes at 2nd Inst. (Fig. 70) include the addition of setae m3ea, emp, and p 7i; the transformation of a1 into a microchaeta, which together with a2 and p3 are displaced close to m2, becoming precursors of the fan setae of the botriothrical complex; and the relocation of m4 closer to a5. At 3rd Inst. the chaetotaxy of area M is completed with the addition of two ciliate microchaetae (Fig. 71). The lateral botriothrical complex is unstable at this instar. All specimens add seta c3 between pm6 and emp, but three additional setae are asymmetrically present, in different combinations, in three of seven specimens examined. The chaetotaxy of the a5 complex remains unstable at 4th Inst.: three specimens have six setae, two have five setae, one has three setae on one side, and one has seven setae on one side. In addition, am6 is a short truncate macrochaeta in two specimens. By 5th Inst. the chaetotaxy is identical to the adult (Fig. 72): all setae in the botriothrical complexes are fan-shaped; am6 is a macrochaeta; and p 7i, and p7 are ciliate, acuminate setae as long as m3.
FIGURES 37–41. 37–40, Mesotergal chaetotaxy; 37, Seira brasiliana ; 38, S. desapercibida ; 39, S. blanca ; 40, S. annulicornis , redrawn from Szeptycki (1979); 41, S. dowlingi , 1st instar metathorax, arrow points at supernumerary seta, asterisk identifies external microsensilla.
FIGURES 42–47. Dorsal metathoracic chaetotaxy; 42–44, Seira dowlingi ; 42, 2nd instar; 43, 3rd instar; 44, Adult; 45, S. blanca ; 46, S. desapercibida , setae m6, m7 and m6p are present but not shown; 47, S. brasiliana , m6p is present but not illustrated.
FIGURES 48–54. 48–50, Seira annulicornis ; 48–50, Metathorax, and alternative interpretations of internal setae homology; 51–54, S. dowlingi , chaetotaxy of first abdominal segment; 51, 1st instar; 52, 2nd instar; 53, 3rd instar; 54, Adult, seta m6 is present but not shown.
FIGURES 55–60. 55–59, Dorsal chaetotaxy of first abdomen; 55, Seira steinmetzi ; 56, S. blanca ; 57, S. brasiliana ; 58, S. desapercibida ; 59, S. annulicornis , redrawn from Szeptycki (1979); 60, S. dowlingi 1st instar, dorsal chaetotaxy of second abdominal segment.
Abdomen IV. In addition to the regular primary and secondary setae, this segment has smooth, tapered, blunt, supplementary setae that often appear thin-walled. As in other segments, primary and secondary setae are structurally indistinguishable in the adult. The supplementary setae appear at 1st Inst. and are, by definition, primary, but their position often varies between specimens and even between sides of the same specimen. The supplementary setae are restricted to the area between the pseudopore and column C, except for ps which is always associated with T7. Supplementary setae are not labeled (except ps) and only their number is mentioned in the following description and discussion.
The chaetotaxy of S. dowlingi is arranged into 7 columns with varying number of setae per column (Fig. 74). Features deserving notice at 1st Inst. are the presence of B1 and the absence of A4 and T1. The primary macrochaetae correspond to B4, B5 and E3. There are 10–11 supplementary setae. The fan-shaped setae associated with the botriothrical complexes and the posterior botriothricum are undifferentiated (Fig. 75). The setae added at 2nd Inst. (Fig. 76) are A3a, A4s, C1p, T1s, T1p, D2a, D3p, E2p, E4p, E4p2, F1p, F3p, three additional setae of uncertain homology postero-external to columns Fe, and five posterior setae. Primary setae B6, C1, F2 and F3 change into macrochaetae. The posterior botriothricum corresponds to seta D3. The setae added at 3rd Inst. (Fig. 77) are Ae7 and Ee10; A3 becomes a macrochaeta, and some setae in the botriothrical complexes change into fans (triangles in Fig. 77). All setae added at 4th Inst. belong to columns external to C, but those on E and F are variable and it is difficult to consistently ascertain their presence or absence in all specimens studied. Setae associated with the bothriotrichial complexes appearing at 4th Inst. (drawing not included) are m, D1p, Pi, and Pe. Setae A4s, B3, T7, E4, E2, and Ee10 are transformed into type two macrochaetae (i. e., acuminate instead of clubbed or truncate). The botriothrical complexes are completed at 5th Inst. (Fig. 79); otherwise, this instar differs from the adult (Fig. 78) only in lacking setae A2a and in the number of macrochaetae in rows E, F, and Fe.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.