
Elminiinae Foster, 1982
|
publication ID |
https://doi.org/ 10.5281/zenodo.275597 |
|
DOI |
https://doi.org/10.5281/zenodo.6196925 |
|
persistent identifier |
https://treatment.plazi.org/id/03D887BC-2164-F55B-FF60-B7D6FC57F8AC |
|
treatment provided by |
Plazi |
|
scientific name |
Elminiinae Foster, 1982 |
| status |
|
Subfamily Elminiinae Foster, 1982 (nom correctum, Buckeridge, 1983a).
Diagnosis: Austrobalanids lacking both internal chitinous stringers and arborescent interlaminate figures.
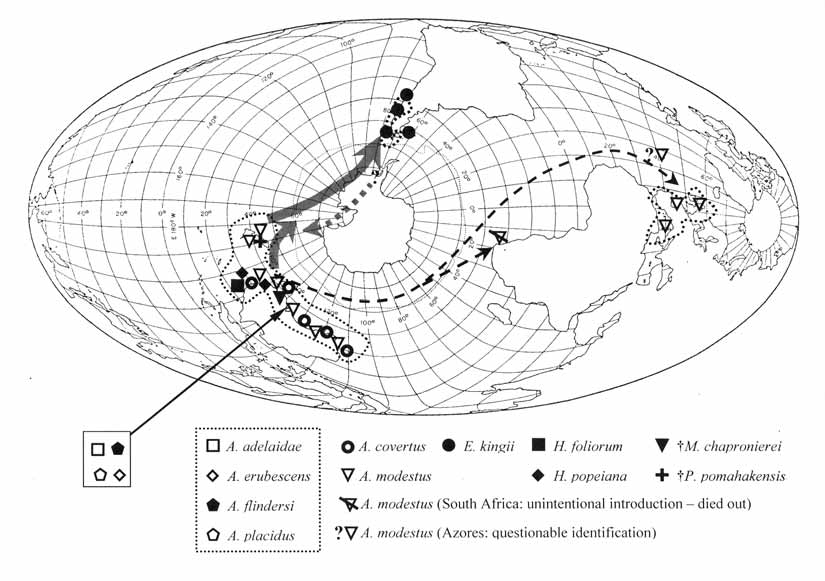
Distribution and age: Littoral, low energy environments. Oligocene to Recent: southeastern Australia, New Zealand, southern Chile around Tierra del Fuegio to southern Argentina and out to the Falkland Islands; introduced into Europe during the 1940s, temporarily to South Africa in the 1950s and questionably introduced to the Azores in the early 1900s ( Figure 1 View FIGURE 1 ).
Remarks: Newman and Ross (1971, copyright 1970), cited by Ross (1970) as “in press”, had discovered chitinous structures in the wall of Elminius simplex similar to those seen in some tetraclitids, and they briefly discussed how this, as well as some previously known characters, suggested tetraclitid rather than balanine affinities. In the discussion of Epopella , the new genus Ross (1970) erected to accommodate Tetraclita (Tetraclita) purpurascens forma breviscutum Broch, 1922 , both Elminius plicatus Darwin, 1854 and E. simplex were transferred to Epopella and thus assigned to the tesseroporan tetraclitids.
As noted in the introduction, Foster (1979) was not aware that Epopella plicata , E. simplex and Epopella breviscutum , had been transferred to the tetraclitid subfamily Austrobalaninae by Newman and Ross (1976). Likely, therefore, he did not feel the need to formalize his new family, the Elminiinae , other than to say, “This species (of Hexaminius ) and of Elminius , seem to constitute a southern hemisphere subfamily as well defined as the northern hemisphere Semibalaninae . I propose the name Elmininae (sic) for this subfamily” ( Foster, 1982a: 22). Foster’s original spelling for the subfamily was “Elmininae” which was derived from the genus Elminius Leach, 1825 , and thus, according to the International Code of Zoological Nomenclature, should have been Elminiinae ( Buckeridge, 1983a; Foster, 1987).
The phylogeny of the elminiines ( Figure 2) draws upon the conclusions reached by Pérez-Losada et al. (2008) plus interpretation of a relatively sparse but nonetheless informative fossil record ( Buckeridge, 1983b, 1984). The main issue with molecular-based reconstructions of phyletic divergence is an assumption that significant changes must occur over considerable time. In Pérez-Losada et al. (2008: 337), it is proposed that the Verrucomorpha-Balanomorpha divergence occurred shortly after the close of the Jurassic c.145 MYBP. However, if the absence of any late Mesozoic balanomorph barnacles is a true reflection of Cretaceous biodiversity (as opposed to an artifact of preservation), and we believe it is, then Protelminius would have arisen from an eight-plated pachylasmatid such as Eolasma only c.50 MYBP.
The absence of Mesozoic balanomorphs is the basis for the broader assessment of balanomorph cirripede divergences as shown in Figure 3 View FIGURE 3 . As the Tetraclitoidea appear first in the late Palaeocene (Thanetian) of the Chatham Islands ( Buckeridge, 1983a), the divergence of the earliest tetraclitoids (e.g. Bathylasma ), from the pachylasmatids, must have occurred in less than 10 million years.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |