
Pujoliclerus pallidus Opitz, 2014
|
publication ID |
https://doi.org/ 10.1649/0010-065X-68.4.727 |
|
publication LSID |
lsid:zoobank.org:pub:CD987054-035A-4A5B-B488-EEA35F632F23 |
|
persistent identifier |
https://treatment.plazi.org/id/03D85669-FFBE-AB1F-FD3F-890DFCA9FCDC |
|
treatment provided by |
Carolina |
|
scientific name |
Pujoliclerus pallidus Opitz |
| status |
sp. nov. |
Pujoliclerus pallidus Opitz , new species ( Figs. 46 View Figs , 66 View Figs , 91 View Figs , 117 View Figs , 127 View Fig )
Holotype. ♂. H O N D U R A S: C o m a y a g u a, 2.8 km NNE Los Planes , 26 May 2002, R. Turnbow ( FSCA).
Paratypes. One specimen. Same label data as the holotype ( RHTC, 1) .
Diagnosis. The yellow body coloration will distinguish this species from its congeners.
Description. Size: Length 6.0 mm; width 1.8 mm. Form: As in Fig. 117 View Figs . Integument: Head mostly yellow: pronotum and elytra yellow; antenna, profemora, pterothorax, and abdomen brown. Head: Eyes wider than vertex (14:8); funicular antennomeres considerably expanded, 6 th antennomere acuminate ( Fig. 46 View Figs ). Thorax: Pronotum quadrate (35:35), lateral tubercle very prominent ( Fig. 66 View Figs ); asetiferous punctures end at elytral apical 1/4, punctures prominent in anterior 2/3; epipleural fold wide in elytral basal half, then gradually diminishing to elytral apex. Abdomen: Tegminal lobes and phallic apex as in Fig. 91 View Figs .
Variation. The available specimens are quite homogeneous.
Natural History. The types were collected during May.
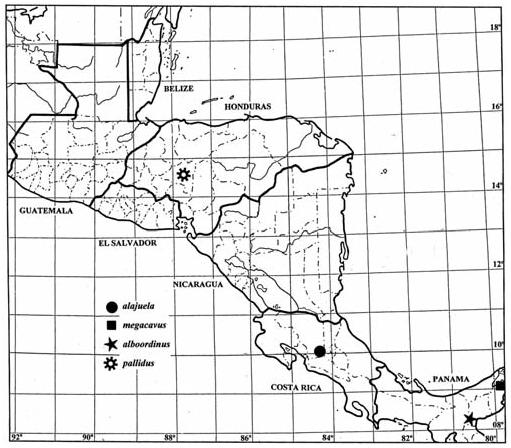
Distribution. Known only from the type locality ( Fig. 127 View Fig ).
Etymology. The specific epithet pallidus (= pale) is a Latin adjective and refers to the predominantly yellow color of this beetle.
EVOLUTIONARY CONSIDERATIONS
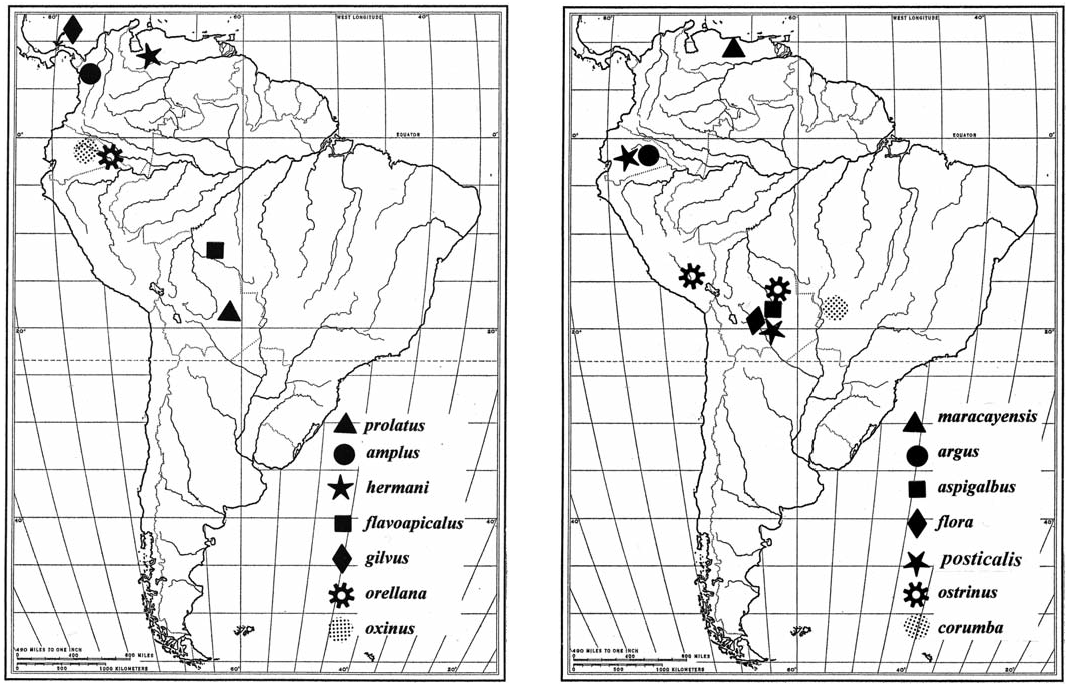
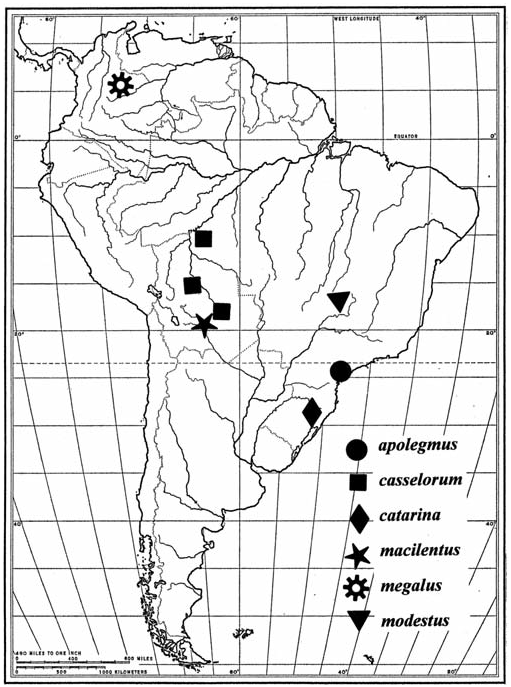
The known distributions of Pujoliclerus species are illustrated in Figs. 123–127 View Fig View Fig View Fig , and their occurrence in refugial forests is noted in Table 2. The
Area of endemism Pujoliclerus species
Central America Nuclear X Insular X X X X
South America Cordillera Occidental X X X X Cordillera Oriental Complex X X X Altiplano Complex X X X X X X X ° X X Amazon Basin X X X ° Brazilian Highlands X X X X X X XX
table indicates the number of species of this genus in endemic regions defined as “clerofaunas” by Opitz (2005), which in essence represent extensions of biogeographic concepts presented by Whitehead and Ball (1997). It is acknowledged that the distribution patterns presented herein are tentative as few specimens of some species became available for study. However, I have assessed that a few biogeographical and evolutionary comments are warranted, if only to set the stage for more refined comments when more material becomes available.
When one views Table 2, it becomes clear that extant Pujoliclerus species are primarily distributed in South American clerofaunas. The lack of diversity in Mexo-America and Central America (sensu Opitz 2005) suggests a fairly recent evolution of the genus with a probable origin in the highlands of South America. It also becomes clear that of the six species-groups recognized herein, four contain species that independently entered Central America, probably after the closure of the Panamanian portal during the early Cenezoic ( White 1986). Ancestors of P. pallidus ( flora species-group) traversed both the Panamanian Portal and the Nicaraguan Depression (Whitehead and Ball 1997) to settle in Honduras. Progenitors of P. alajuela and P. megacavus ( alajuela speciesgroup), P. gilvus ( oxinus species-group), and P. alboordinus ( casselorum species-group) dispersed northward across the portal to settle in central Costa Rica and lower Panama. Twenty-four species originated and remained in the highlands of South America. To explain the historical events that produced the extant distributions of the South American species, I rely on the concept of the center of origin and dispersal resulting from historical orogenic and climatic events.
For example, Pujoliclerus speciation events in the Cordillera Occidental and the Cordillera Oriental would have been greatly influenced by the changing vegetative character of these highlands. Such vegetation would have been greatly affected by the cooling and drying effect of glacial periods ( Simpson 1974) and by the uplift activities of the Miocene to produce montane ranges such as the northern Andes. In general, Cleridae are most abundantly collected in high montane oak/pine forests ( Ekis 1977; Opitz 2006, 2007, 2008).
The sister genera Cregya and Pujoliclerus are predominantly distributed in South America. This suggests a South American origin for the ancestral stock of these taxa. The progenitor of Cregya - Pujoliclerus (ancestor A) would have evolved an obliquely positioned epipleural fold and a bipartite phallic apex. This progenitor would have diverged to produce ancestor B and a lineage that led to ancestor C, and in the process the funicular antennomeres began the progression towards an explanate condition. Ancestor B eventually generated a line in which body length was increased as seen in the extant P. maracayensis . Ancestor C retained the short body form and evolved subsecuriform terminal palpomeres. Moreover, Ancestor C produced the flora species-group whose species developed abbreviated elytral punctation. Ancestor C also led to ancestor D and towards a transformation that led to bicolorous elytra and eventually developed the progenitor of the flavolimabatus species-group ancestor. Eventually, antennomere 8 became greatly lengthened in the evolutionary line that generated the flavolimbatus species-group. Then ancestor E taxa developed a line that evolved the oxinus species-group and a second line that generated ancestor F. Ancestor F produced an eventual evolutionary bifurcation in which the epipleural fold became yellow ( casselorum species-group) and the pronotum became partially reddish in the line that traversed into Insular Central America ( alajuela species-group).





| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.