
Calea × parviantha V.R.Bueno & G.Heiden, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.579.3.1 |
|
DOI |
https://doi.org/10.5281/zenodo.7555258 |
|
persistent identifier |
https://treatment.plazi.org/id/03D8054C-BF65-FF9B-FFEE-066BFBDFFC28 |
|
treatment provided by |
Plazi |
|
scientific name |
Calea × parviantha V.R.Bueno & G.Heiden |
| status |
sp. nov. |
Calea × parviantha V.R.Bueno & G.Heiden View in CoL , nothosp. nov. ( Fig. 4 View FIGURE 4 , 5 View FIGURE 5 )
Calea × parviantha differs from C. parvifolia by the striate stems (vs. corrugate stems), hispid or hispidulous or glabrous stems (vs. puberulous or puberulent), and coriaceous leaves (vs. chartaceous leaves); and from C. triantha by the rounded base leaf, sometimes subcordate (vs. cordate), concave paleae (vs. conduplicate), and pappus scales 1.1–2.7 mm long (vs. 0.8–1.3 mm).
Type:— BRAZIL. Paraná, Jaguariaíva, campo próximo à ponte do Rio das Mortes, PR-151, 16 December 1991, A. C. Cervi et al. 3546 (holotype: MBM!, isotypes: CGMS!, CPAP!, UPCB!) .
Shrubs, 0.2–1.2 m tall, stems castaneous, cylindrical, striate, hispid, often sparsely hispid, rarely densely hispid or hispidulous or glabrous, rarely glandular-punctate, internodes 5.4–32.7 mm long. Leaves decussate, petiole 0.5–2.2 mm long; blades olivaceous or dark green adaxially, concolorous, sometimes slightly discolorous or discolorous, coriaceous, 0.78–2.57 × 0.42–1.59 cm, ovate or narrow ovate, rarely wide ovate, base rounded, sometimes subcordate, apex acute, rarely attenuate or obtuse, venation basal acrodromous and reticulate or basal acrodromous, margins revolute, often slightly revolute, entire or pauciserrate, sometimes serrate, 0–12 (–13–14) teeth, 0.3–1.8 mm long, from the base to middle, sometimes absent or just in the middle, rarely along the margins, abaxial surface hispidulous, often glabrous, sometimes hispid, veins hispid to hispidulous or hispidulous, sometimes glabrous, densely glandular-punctate, often glandular-punctate, adaxial surface scabridulous, often sparsely scabridulous with margin scabridulous, sometimes glabrous to scabridulous, glands absent. Capitulescence cymose dichasial, sometimes capitulum solitary, profusely branched or branched, axis 0–14.56 (–30.3) cm long, branching to 2–3 rd ramifications, rarely 4–5 th ramifications, 2–18 secondary stems, peduncle 0.08–3.55 (–3.9–4.16) cm long, hispid, rarely glandular-punctate. Capitulum heterogamous, radiate, involucrum campanulate, 5.8–9.7 × 6.8–13.6 mm, 5-seriate; phyllaries conspicuously or inconspicuously striated; two outermost series of phyllaries olivaceous, sometimes olivaceous with base yellowish green, foliaceous, sometimes foliaceous with base scarious, margins entire, sometimes serrate, abaxial surface hispid to hispidulous, sometimes glabrous or hispidulous, densely glandular-punctate or glandular-punctate; first series 4–6 striate, blades 6.6–10.2 × (3–3.3–) 3.9–4.7 (–5.5–5.7) mm, wide elliptic or narrow obovate, sometimes elliptic, rarely oblanceolate, apex acute or obtuse, half revolute, often revolute; second series 4–8 striate; blades 7–10.4 × 3.1–5.7 (–7.5) mm, obovate or wide elliptic or panduriform, apex obtuse, often acute, revolute, rarely half revolute; third series yellowish green to olivaceous, apex foliaceous, sometimes scarious, 6–10 striate, blades 5.5–9.7 × 2.7–4.8 mm, oblong or narrow ovate, sometimes narrow obovate, margins entire, apex obtuse, sometimes rounded, rarely acuminate, revolute or flat; abaxial surface sparsely hispid at apex or pilose at apex, sometimes glabrous, densely to moderately glandular-punctate at apex, sparsely to densely glandular-punctate; two innermost series yellowish green to light yellow or yellowish green, scarious, glabrous, sometimes glandular-punctate at apex, fourth series 8–15 striate, blades 7.2–9.9 × 3.3–4.3 mm, oblong, sometimes narrow obovate, margins entire, sometimes erose at apex, apex rounded; fifth series 10–15 striate, blades 7.4–10.9 × 2.3–3.9 mm, narrow oblong or oblanceolate, sometimes narrow elliptic, margins entire, apex rounded, sometimes obtuse; receptacle slightly convex, holopaleaceous; paleae light yellow, concave, 7–9 mm long, narrow elliptic or narrow oblong, sometimes oblanceolate, apex long acuminate. Ray florets 8–15, (10.2–) 12–15.7 (–19.8) mm long, pistillate, corolla liguliform, (8.3–) 9.3–14.2 (–16.4) mm long, yellow, tube 1.5–3.8 mm long, limb 5.9–12.6 × (1.8–2.3–) 3.2–4.6 mm, narrow obovate, often narrow elliptic or narrow oblong or oblanceolate or elliptic, apex rounded, sometimes 2–4 lobulate, rarely obtuse, 5–7 veins, abaxial surface densely glandular-punctate, sometimes glandular-punctate, adaxial surface glabrous; style arms yellow, 0.7–1.5 mm long. Disc florets 30–55, 7–10 mm long, bisexual, corolla tubular, 4.4–7 mm long, yellow, tube 1.3–2.6 mm long, lobes 1–1.7 (–2.1) mm long, sparsely glandular-punctate or glands absent, rarely glandular-punctate; anthers light yellow, (2.3–) 2.5–2.9 mm long, apical appendages ovate; style arms yellow, 0.8–1.2 mm long, linear. Cypselae blackish, 2.2–3.2 (–3.6) mm long, prismatic, ray cypselae 3-angled, disc cypselae 4-angled, glabrous to pilose or densely pilose or sparsely pilose to pilose, ribs densely pilose, often pilose; pappus monotypic, often bitypic, scales monolength, sometimes bilength, 8–15 (1–2 distinctly longer, when present), free, 1.1–2.2 mm long, longer scales 1.6–3 mm long, when present, oblong or oblanceolate, sometimes narrow elliptic, apex obtuse or rounded, margins entire, erose at apex.
Distribution, Habitat and Predicted Area of Occurrence:— Calea × parviantha occurs in two Brazilian states: S„o Paulo and Paraná ( Fig. 2 View FIGURE 2 ). A total of eight municipalities have occurrences of this new nothospecies, two in S„o Paulo and six in Paraná. In addition, Ponta Grossa (municipality in Paraná) has an occurrence only recorded by photos (Pricila Just, pers. comm.). The new nothospecies occurs in areas of grasslands with rocks outcrops and riverine vegetation along the contact zones of high elevation tropical grasslands of the Atlantic Forest domain and the southernmost limits of open grasslands and savannas from the Cerrado domain. In the Jaguariaíva sites, C. × parviantha is more commonly found than its parental taxa ( Fig. 2 View FIGURE 2 ), suggesting that hybrid populations are probably replacing them in this area.
According to the niche modelling, the new nothospecies potentially occurs in open areas where the two parental species are allopatric from the center-south of S„o Paulo to the south of Paraná ( Fig. 3 View FIGURE 3 ). However, due to its hybrid nature, it is more likely that the distribution of the nothospecies is restricted from Itararé, S„o Paulo, to Lapa, Paraná, where the parental species co-occurrence is likely and the contacts between Atlantic Forest and Cerrado are more common ( Fig. 2 View FIGURE 2 ).
Informal Conservation Status:— Calea × parviantha could be considered Endangered (EN). Traditionally, hybrids are not evaluated for their conservation status, but as this hybrid has different collections over almost a century, it is probably a stable hybrid and therefore this status was evaluated. The GeoCAT ( Bachman et al. 2011) analyses generated an EOO of 5,000 km ² and AOO of 48 km ², both values suggesting EN status. C. × parviantha also can be considered EN due the criteria ( IUCN 2019): B1B2; C2(i); D. The new nothospecies mostly occurs out of protected areas, except for three occurrences in Ecologic Station of Itapeva (Itapeva, S„o Paulo), Ecological Park of Barreira (Itararé, S„o Paulo) and probable occurrence in Cerrado State Park (Jaguariaíva, Paraná). The few records in protected areas, the AOO and EOO values and the above-mentioned IUCN criteria are strong subsidies for classifying Calea × parviantha as Endangered.
Etymology:—The nothospecific epithet “parviantha” is proposed based on the merging of the specific epithets of the putative parental species: Calea parvifolia and C. triantha .
Phenology:—Most of the collected specimens were found flowering and fruiting from December to April, with one outlier record flowering in August.
Taxonomic comments and support analysis: —Most of the specimens of Calea × parviantha were misidentified as C. myrtifolia , mainly due to the ovate leaves bigger than the leaves of C. parvifolia . Studies of specimens labeled as C. myrtifolia from S„o Paulo and Paraná evidenced that some specimens were the hereby described C. grandiflora and the remaining a putative hybrid between C. parvifolia and C. triantha . The main morphological characteristics that indicated these specimens as a new hybrid species were the combination of reticulate venation and hispid indumentum—features absent in C. myrtifolia , but present in C. parvifolia and C. triantha , respectively. Therefore, based on the evaluation of all the available specimens previously labeled as C. myrtifolia we can currently confirm that this species is endemic to the state of Minas Gerais in Southeastern Brazil.
Calea × parviantha shares with C. parvifolia the rounded leaf base, occasionally subcordate, elliptic blade of first series of phyllaries, concave paleae of receptacle, tube of disc floret 1.3–2.6 mm long, most of cypselae with glabrous to pilose face and densely pilose ribs, and pappus monotypic, often bitypic in the same plant. They can be distinguished by the striate stems (vs. furrowed stems), hispid or hispidulous or glabrous stems (vs. puberulous or puberulent), glands absent in stems, rarely glandular-punctate (vs. densely glandular-punctate, sometimes glandular-punctate), coriaceous leaves (vs. chartaceous), revolute margin, sometimes slightly revolute (vs. flat), commonly hispidulous abaxial surface (vs. commonly glabrous), revolute apex in second series of phyllaries (vs. flat), and always entire margin in third series of phyllaries (vs. mostly serrate, rarely entire).
Calea × parviantha has striate stems and coriaceous leaves in common with C. triantha . In addition, these species commonly have hispid and eglandular stems, hispid peduncle, hispid to hispidulous outer phyllaries, and rounded apex of ray florets. The nothospecies differs from this parental species by the commonly ovate or narrow ovate leaves (vs. very wide ovate, sometimes wide ovate, rarely ovate), rounded base leaf, sometimes subcordate (vs. cordate), usually acute apex (vs. usually obtuse apex), concave paleae (vs. conduplicate), pappus monotypic, often bitypic (vs. always monotypic), monolength pappus scales, often monolength and bilength (vs. only monolength), and pappus scales 1.1–2.7 mm long (vs. 0.8–1.3 mm).
The characteristics that the nothospecies presents that are intermediate or that overlap with those of the parental species are listed in Table 1 View TABLE 1 . The PCA results corroborate the hypothesis on the hybrid origin of C. × parviantha , all characters listed in Table 2 View TABLE 2 contributed to the separation of taxa in the PCA analysis. The multivariate analyses plot ( Fig. 6 View FIGURE 6 ) evidences that the morphological data groups the specimens in distinct groups of parental species and the nothospecies with intermediate values.
Examining all the specimens, a gradation of characters was seen in the different populations, but within these, some morphological standardization is seen among the specimens, mainly in those found in the municipality of Jaguariaíva and region, Paraná state. All hybrid populations are in contact with populations of the parental species, the nothospecies populations further south of the distribution are more than the Jaguariaíva populations (north of the nothospecies distribution). We believe that due to all the evidence presented here it is a nothospecies; however, more molecular studies would be needed to corroborate this hybrid status.
Paratypes:— BRAZIL. Paraná: Balsa nova, Ponte dos Arcos , 07 December 2005, C . Kozera 2688 ( MBM); Jaguariaíva, campo próximo à ponte do Rio das Mortes - PR 151, 16 December 1991, C . Kozera & O . P . Kozera 2688 ( MBM); Castro , entre Castro e Ponta Grossa, 1950, J . Vidal 3-151 ( R); Jaguariaíva , campos, 05 February 1910, P . Dusén 9187 ( S); campos, 26 December 1914, P . Dusén 16195 ( MO, US) ; rio das Mortes , 02 February 1995, J . R . Stehmann & J . Semir 2169 ( BHCB, UEC); montanha, 07 April 2007, F . Bueno s.n. ( RB); Palmeira, fazenda Santa Rita , 24 January 1990, L . T . Dombrowski et al. 14258 ( MBM); Sengés, mata ciliar na margem esquerda do Rio Funil , 08 April 1995, Ribeiro Jr. et al. 04 ( FUEL, SP, UEC); Tibagi , Parque Estadual Canyon Guartelá , 10 February 1997, V . F . Kinupp et al. 273 ( FUEL, UEPG). S „o Paulo: Itapeva, Estaç„o Ecológica de Itapeva , 18 August 1995, V . C . Souza et al. 8732 ( ESA, SP, SPF); 27 January 2009, J . B . Baitello & C . R . Lima 2236 ( MBM, SPSF); Estaç „o Ecológica de Itapeva, trilha da Vila, 26 March 2008, J. B. Baitello et al. 2051 ( SPSF, UEC); Itararé, Gruta da Barreira, 04 December 1984, C. Muller et al. 5994 ( FUEL, UEC); próximo à entrada da Fazenda S „o Nicolau, 21 April 1993, V . C . Souza et al. 3981 ( ESA) .
Additional specimens examined:— Calea parvifolia — BRAZIL. Paraná: Arapoti, Estrada para Ventania na Barra do Rio Perdizes com o Rio das Cinzas , beira da estrada, 03 March 2003, M. A. G . Magenta 558 ( MO, SPF); Balsa Nova, Ponte dos Arcos, afloramento rochoso próximo a Mata de Pinus , 02 April 2019, A. L . Christ et al. 646 ( ICN); Bocaiúva do Sul, Serra da Bocaina , 20 April 1998, J. M . Silva et al. 2331 ( MBM); Ponta Grossa, Furnas de Itaiacoca , 20 August 2014, J . Cordeiro et al. 4705 ( MBM); Tibagi, Parque Estadual do Guartelá , Rio Iapó , 01 September , S. R . Ziller 1578 ( MBM). S „o Paulo: Itapeva, Estaç„o Experimental , 03 November 2009, R . Cielo-Filho et al. 896 ( SPSF) .
Calea triantha — BRAZIL. Minas Gerais: Poços de Caldas, Campo do Saco , 12 Feb 1965, M . Emmerich 2381 ( RB) . Paraná, Campo Largo, fazenda Thalia , 08 March 1998, W . Amaral 22 ( MBM); Ventania, fazenda Santa Inês, 11 February 2005, D. A . Estevan et al. 531 ( HUFU) . Santa Catarina, Lages, salto Caveira , 29 January 2015, R . Mello-Silva 3765 ( SPF); Rio Negrinho, salto do rio Antinha , 23 December 2018, C . Ribeiro et al. 313 ( ICN). S „o Paulo, Campos do Jord„o, vale do rio Coxim , areal, 17 March 1964, J . Correa Gomes Jr. 1620 ( SP) .
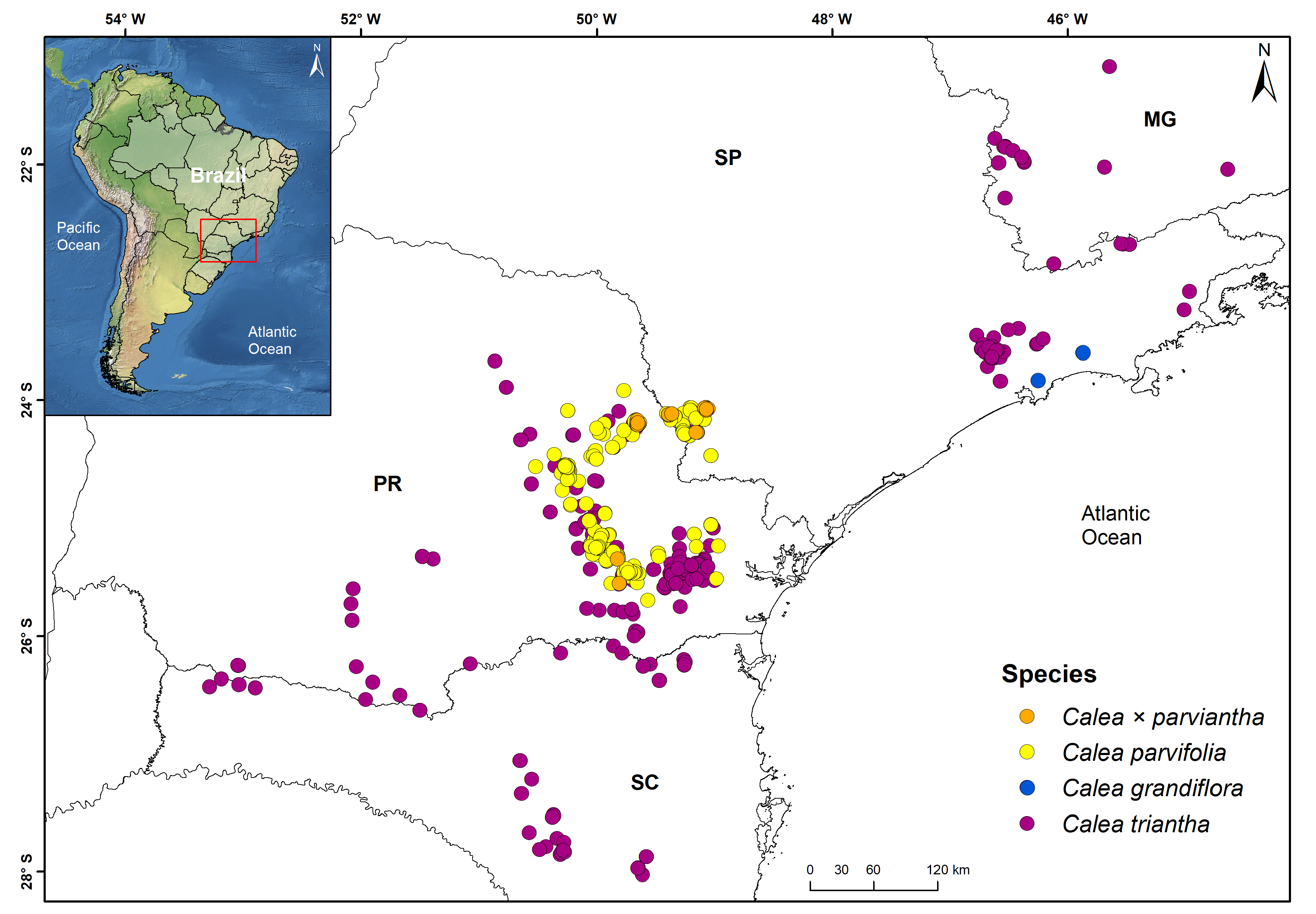




TABLE 1. Vegetative and reproductive informative characteristics measured for the Principal Component Analysis performed with Calea × parviantha and its parental species C. parvifolia and C. triantha (Neurolaeneae, Asteraceae).
| C. parvifolia | C. × parviantha | C. triantha | |
|---|---|---|---|
| Venation | reticulate | basal acrodromous and reticulate or basal acrodromous | basal acrodromous |
| Leaf margin | entire | entire or pauciserrate, rarely serrate | serrate |
| Glands in peduncle | densely punctate | eglandular, rarely glandular-punctate | eglandular |
| Margin of outer phyllaries | pauciserrate, rarely entire | entire, sometimes pauciserrate | entire |
| Third series of phyllaries (mm) | 8.8–10.3 | 5.5–9.7 | 5.1–7.9 |
| Fifth series of phyllaries (mm) | (8.1) 9–10.2 | 7.4–10.9 | 6.6–8.2 (9.3) |
| Paleae | narrow elliptic | narrow elliptic or narrow oblong, sometimes oblanceolate | oblong |
| Disc floret length (mm) | 8.5–11.6 | 7–10 | 6.1–8.4 |
| Glands on disk floret corolla | glandular-punctate | sparsely glandular-punctate or eglandular, rarely glandular-punctate | eglandular |
| Length of pappus scales | monotypic, often bitypic | monotypic, often bitypic | monotypic |
| Length of pappus scales | monolength or bilength | monolength, sometimes bilength | monolength |
TABLE 2. Intermediate or overlapped morphological features of Calea × parviantha and its parents: C. parvifolia and C. triantha (Neurolaeneae, Asteraceae).
| Informative Characters | ||
|---|---|---|
| Internodes | Measurement | Length |
| Petiole | Measurement | Length |
| Measurement | Length | |
| Vegetative | Measurement | Wide |
| Leaf | Characterization | Shape of apex |
| Characterization | Types of venation | |
| Characterization | Types of margins | |
| Peduncle | Measurement | Length |
| Involucre | Measurement Measurement | Length Wide |
| Striate phyllaries | Characterization Counting | Stripes conspicuous or inconspicuous Number |
| Second order of phyllaries | Characterization | Shape |
| Measurement | Length | |
| Measurement | Wide | |
| Third order of phyllaries | Characterization Characterization | Shape Apex |
| Characterization | Glandular or eglandular | |
| Characterization | Margins | |
| Measurement | Length | |
| Fourth order of phyllaries | Measurement | Wide |
| Characterization | Glandular or eglandular | |
| Fifth order of phyllaries | Measurement Measurement | Length Wide |
| Paleae | Measurement Characterization | Length Apex |
| Reproductive | Counting | Number |
| Measurement | Length of corolla | |
| Measurement | Length of limb | |
| Ray florets | Characterization | Shape of apex |
| Counting | Veins in corolla | |
| Characterization | Glandular or eglandular | |
| Measurement | Length of style | |
| Measurement | Length | |
| Measurement | Length of Corolla | |
| Disc florets | Measurement | Length of Tube |
| Measurement | Length of Limb | |
| Characterization | Glandular or eglandular | |
| Anthers | Measurement | Length |
| Style | Measurement | Length |
| Cypselae | Measurement Characterization | Length Indumentum of surface |
| Measurement | Length | |
| Pappus scales | Characterization | Shape |
| Characterization | Apex | |
| A |
Harvard University - Arnold Arboretum |
| C |
University of Copenhagen |
| MBM |
San Jose State University, Museum of Birds and Mammals |
| CGMS |
Universidade Federal de Mato Grosso do Sul |
| CPAP |
Centro de Pesquisas Agropecuarias do Tropico Umido |
| UPCB |
Universidade Federal do Paraná |
| O |
Botanical Museum - University of Oslo |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| J |
University of the Witwatersrand |
| R |
Departamento de Geologia, Universidad de Chile |
| S |
Department of Botany, Swedish Museum of Natural History |
| MO |
Missouri Botanical Garden |
| BHCB |
Universidade Federal de Minas Gerais |
| UEC |
Universidade Estadual de Campinas |
| F |
Field Museum of Natural History, Botany Department |
| RB |
Jardim Botânico do Rio de Janeiro |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| T |
Tavera, Department of Geology and Geophysics |
| FUEL |
Universidade Estadual de Londrina |
| SP |
Instituto de Botânica |
| V |
Royal British Columbia Museum - Herbarium |
| ESA |
Universidade de São Paulo |
| SPF |
Universidade de São Paulo |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| SPSF |
Instituto Florestal |
| M |
Botanische Staatssammlung München |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| ICN |
Instituto de Ciencias Naturales, Museo de Historia Natural |
| W |
Naturhistorisches Museum Wien |
| HUFU |
Universidade Federal de Uberlândia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.