
Vitreorana parvula ( Boulenger, 1895 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5415.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:1EB7FBA8-5DE3-4051-AC91-8E67D538DE89 |
|
DOI |
https://doi.org/10.5281/zenodo.10707520 |
|
persistent identifier |
https://treatment.plazi.org/id/03D73848-FFA0-FFFF-529F-946AFDDD7908 |
|
treatment provided by |
Plazi |
|
scientific name |
Vitreorana parvula ( Boulenger, 1895 ) |
| status |
|
Vitreorana parvula ( Boulenger, 1895) View in CoL
Figs. 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7
Hylella parvula Boulenger, 1895: 646 View in CoL ( Fig. 6 View FIGURE 6 ).
Hyla (Hylella) uranoscopa M̧ller, 1924: 234, new synonym ( Fig. 7 View FIGURE 7 ).
Centrolenella uranoscopa Noble, 1926:18 View in CoL .
Cochranella albotunica Taylor & Cochran, 1953: 1648 View in CoL .
Cochranella dubia Taylor & Cochran, 1953: 1653 View in CoL .
Cochranella lutzorum Taylor & Cochran, 1953: 1638 View in CoL .
Cochranella parvula Taylor & Cochran, 1953: 1650 View in CoL .
Cochranella uranoscopa Taylor & Cochran, 1953: 1635 View in CoL .
Cochranella vanzolinii Taylor & Cochran, 1953: 1646 View in CoL .
Centrolenella albotunica Duellman, 1977: 63 View in CoL .
Centrolenella dubia Duellman, 1977: 188 View in CoL .
Centrolenella lutzorum Duellman, 1977: 191 View in CoL .
Centrolenella parvula Duellman, 1977: 192 View in CoL .
Centrolenella uranoscopa Duellman, 1977: 196 View in CoL .
Centrolenella vanzolinii Duellman, 1977: 196 View in CoL .
Hyalinobatrachium parvulum Ruiz-Carranza & Lynch, 1991: 24 View in CoL .
Hyalinobatrachium uranoscopum Ruiz-Carranza & Lynch, 1991: 25 View in CoL .
Vitreorana uranoscopa Guayasamin et al., 2009: 39 View in CoL .
Lectotype. BMNH 1947.2 .12.67, adult male from municipality of Lages , state of Santa Catarina, Brazil.
Topotypes. Nine specimens, all adult males: MCP 15451–59 View Materials . Collected by Victor M. Zucchetti and Marco Odicio-Iglesias on 09, 12–13 January, 2021, in the Parque Natural Municipal Jo„o José Theodoro da Costa Neto (PARNAMUL), municipality of Lages, state of Santa Catarina, Brazil (-27.7906, -50.3463, 923 m a.s.l.) GoogleMaps .
Synonym justification. The lectotype is an adult male, based on the presence of vocal slits, nuptial pads, and testicles ( Fig. 6 View FIGURE 6 ). The ventral skin is cut transversally so that the coelom can be accessed. Heyer (1985) diagnosed V. parvula from V. uranoscopa using the following characters, states of the latter in parentheses: (1) upper lip ridge absent (present and enameled); (2) snout slightly truncate in profile (spatulate shaped); (3) cloacal ornamentation formed by granular set patch (pair of well-developed horizontal flaps below and to either side of the cloaca); (4) ulnar, tarsal, and foot external folds absent (present, enameled, and scalloped). We argue that these differences result from preservation artifacts and intraspecific variation. Although the individual is not terribly preserved, it is obvious that it has suffered desiccation and compression ( Fig. 6 View FIGURE 6 ). The skin is generally loose over the muscles forming bags of skin, which is a typical result of hydration of a desiccated specimen, while the texture of the ventral skin surfaces is too pronounced compared to any other well-preserved Vitreorana ( Fig. 6 View FIGURE 6 ), which is a typical result of skin shrinking due to water loss. We argue that this cycle (or cycles) of skin shrinking, and expansion has altered subtle dermal characters such as small ulnar, tarsal, and lip folds. Furthermore, these folds are not uniformly conspicuous among all individuals of V. parvula and V. uranoscopa ; some specimens show rather thin and low folds while others show very evident ones. It is likely that the lectotype had weak dermal folds to begin with that have become inappreciable after more than a century in preservative. Regarding the snout shape in profile, Heyer (1985) already argued that this seems to be a preservation artifact. We agree with him.
Besides the evidence outlined above compatible with desiccation, which could cause some changes in head shape, the specimen seems to have experienced a compressive force. This is evident from observing the head in dorsal view, where the left side is more protruding than the right ( Fig. 6 View FIGURE 6 ). A compressive force could make the soft tissues of the snout become shorter and more elevated; thus, giving the impression of a truncated snout in profile. As with dermal folds mentioned above, certain variation exists within well-preserved specimens of V. uranoscopa , and the snout in profile varies from gently sloping to markedly spatulate. Heyer (1985) also points out that “tympani are completely hidden” in the lectotype, while Boulenger (1895), in the original description, describes the tympanum as distinct with about one-third of the eye diameter. The skin of tympanum is very sensitive to preservation conditions and a poorly preserved specimen could result in an under- or over-expression of it. This would explain why Boulenger (1895) observed a tympanum, while Heyer (1985) and we do not.
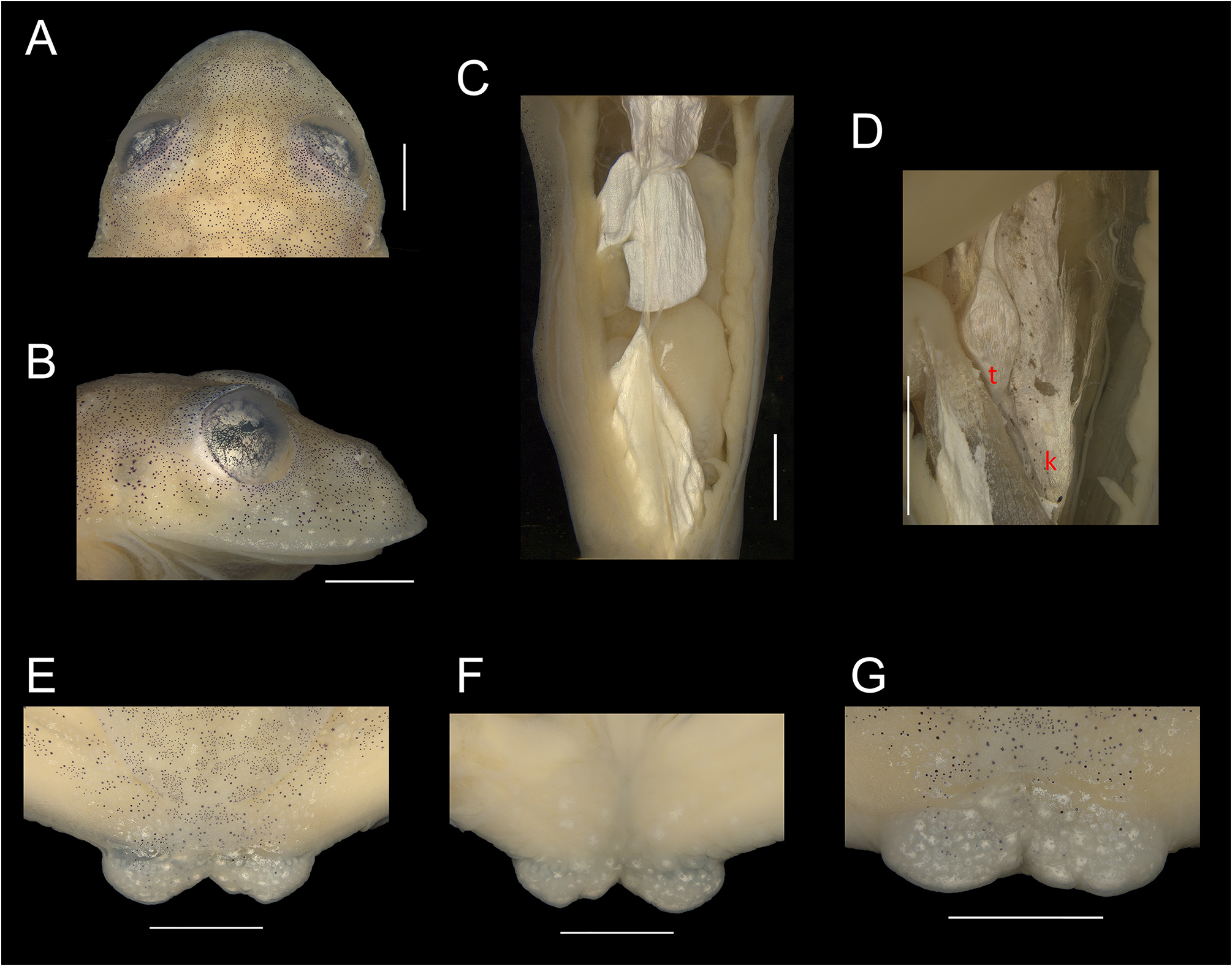
As Heyer (1985) and many after him have recognized (e.g., Cisneros-Heredia & McDiarmid 2007), the accumulation of iridophores formed by guanine crystals in the integument of glassfrogs that gives the characteristic shiny white enameled aspect to structures such as ulnar and tarsal folds, cloacal ornamentation or lip border are easily lost in ethanol preserved specimens, particularly after decades. This seems to be the case with the lectotype of V. parvula that with more than 125 years in ethanol completely lacks integumental iridophores. Finally, the cloacal ornamentation of the lectotype is similar in shape, structure, and position to that of all other topotypic individuals as well as that observed in V. uranoscopa . As shown in Figs. 1E–G View FIGURE 1 and 6D View FIGURE 6 , it is composed by two horizontal flaps of skin. An additional distinction is the small size of the lectotype (SVL = 17.6 mm), which falls outside the variation of other V. parvula (19.4–25.9 mm, Table 1 View TABLE 1 ). However, this could be explained due to preservation artifacts related to desiccation and compression and/or a small adult male.
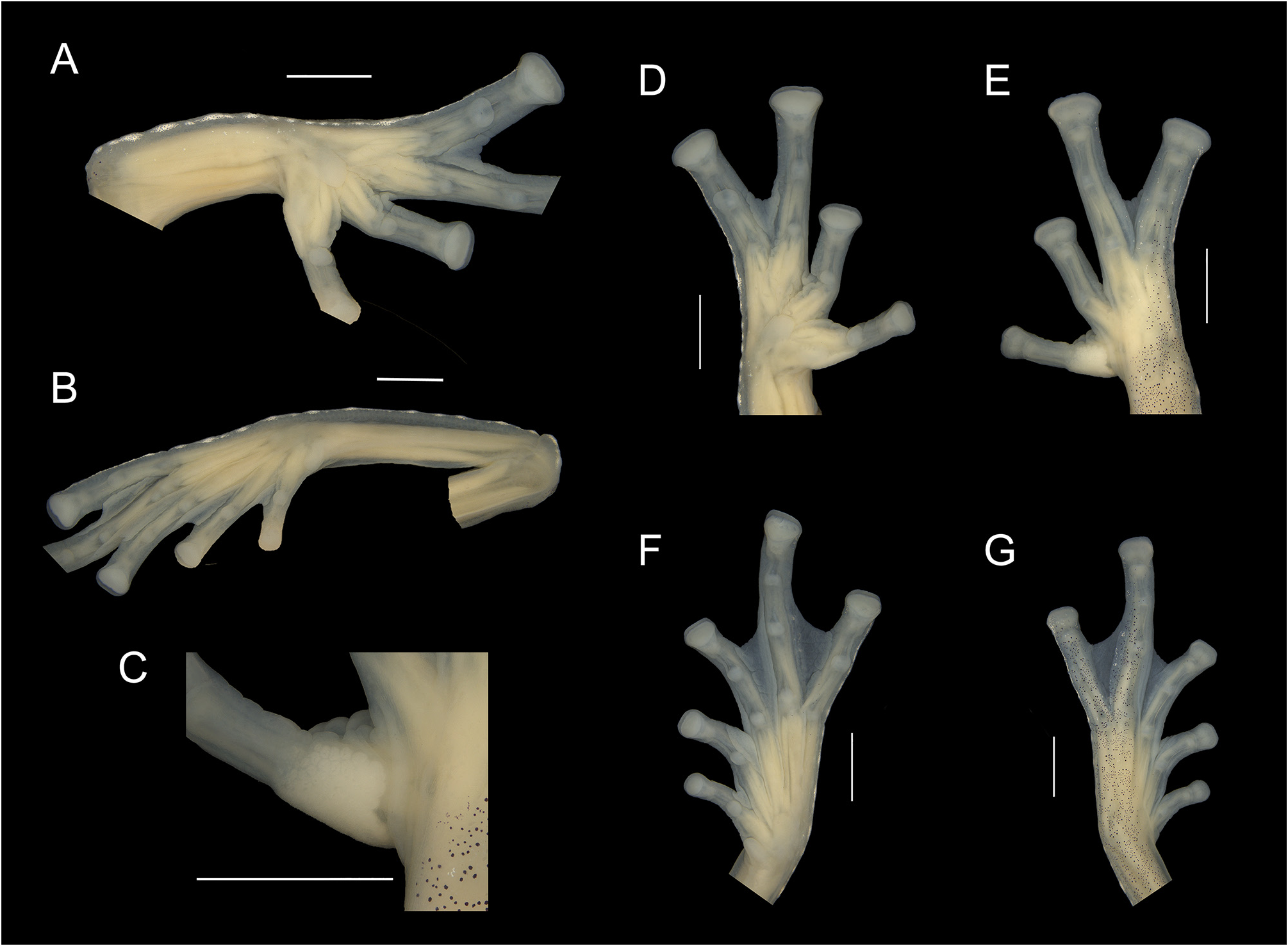
Characterization. (1) absence of teeth in dentigerous process of vomer; (2) snout subovoid to rounded in dorsal view and sloping to strongly sloping in profile; (3) tympanum evident, directed upward, generally rounded, small to moderate in size, its diameter 26.0–36.2% the diameter of eye; tympanic annulus visible except for upper border covered by a weak supratympanic fold extending from behind the eye until the shoulder; tympanic membrane differentiated and partially translucent, enameled in the lower portion or completely with a thick layer of iridophores and partially or completely pigmented with melanophores as surrounding skin; (4) dorsal skin texture shagreen; spicules present; (5) ventral surface areolate; cloacal ornamentation consisting of a skin sheath above the cloaca, an aggregation of medium to large thickened tubercles, which are partially fused forming a pair prominences in each side of cloacal opening, giving an aspect of flesh flaps; (6) parietal peritoneum completely translucent (condition P0), pericardium and hepatic, urinary blader, kidney (dorsal surface only), and gonad peritonea white; gastrointestinal peritoneum translucent (condition V4); (7) bulbous liver, covered by white peritoneum (condition H2); (8) humeral spine absent (crista ventralis not protruding); (9) webbing absent between fingers II and III, reduced between fingers III and IV, and moderate between fingers IV and V, webbing formula: III (2–2 +) (32/3 – 31/2) IV (21/3 –2 +) (2 + –2) V; (10) webbing between toes moderate, webbing formula: I (2 –– 1 +) (2 + –2) II (1 + –1 –) (21/2 –2 +) III (11/3 –1 –) (22/3 –2 –) IV (22/3 –2 +) (11/2 –1) V; (11) evident, crenulated and enameled outer and tarsal folds, extending from the base of Finger V and Toe V to the elbow and ankle, respectively, formed by thickened and enameled tubercles; inner tarsal fold normally present; a row of enameled tubercles, fold-like, on the external side of the tibia; (12) concealed prepollex; nuptial pad present in males, formed by densely packed glands, forming a defined elongate pad, pale colored, on dorsomedial surface of Finger I, individual glands can occur around the pad (Type I); (13) Finger II shorter or about equal to Finger III (Finger II length 86.7–99.3% of Finger III); (14) disc of Finger IV moderate, its width 46–58% of eye diameter; (15) in life, dorsum leaf green with dark punctuations forming reticulations, presence of enameled iridophores along the dorsum, also covering tympanum, upper lip, and borders of nostril; bones green; (16) in preservative, background coloration cream with minute melanophores, and some star-shaped, clustered in arrangement, forming a slightly to strongly reticulated pattern; (17) in life, iris yellowish white with dark brown reticulations and occurrence of a circumpupillary ring; in preservative, iris silvery white with dark grey reticulations; (18) hands and feet yellowish green, sometimes with reddish hues on tips; melanophores usually present in Finger V and toes IV and V; (19) males call from upper side of leaves; call resembling a “trii” ( Duarte-Marin et al. 2022), emitted singly or in groups of 2–9 calls (3.8 ± 2.5), with calls consisting of 1–5 pulses (3.1 ± 0.8) and call duration of 4.3–43.2 ms (24.1 ± 6.5 ms); mean dominant frequency at 4583 ± 251 Hz; call envelope generally triangular pointed right; (20) males fight on leaves, often hanging from leaves in an amplexus-like position; (21) eggs deposited on the upper side of leaves; clutch size of 25– 27 eggs involved by a jelly-rich matrix, presenting empty capsules; early-stage clutches with general cream coloration and light brown animal pole; late-stage clutches with light brown coloration presenting reddish hues and yellowish cream vitellus; short-term maternal care unknown; long-term parental care absent; (22) tadpole snout apparently rounded; spiracle sinistral, located posteriorly; tail long, low; mouth anteroventral; labial tooth row formula 1(1-1)/2, 1(1-1)/1(1-1) or 1(1-1)/3; color light or tan in preservative, irregularly dispersed pigment cells, denser dorsally, that can be uniform on snout region ( Heyer 1985); lateral ridge papilla on buccal roof trifurcated; presence of 8–9 lateral buccal floor arena papillae ( Dias et al. 2020); (23) males with minute to small body size, SVL 20.9–24.4 mm (n = 9); (24) small testis size (about to or less than 1/3 the length of the kidney). Osteological characters described in Zucchetti et al. (2023).
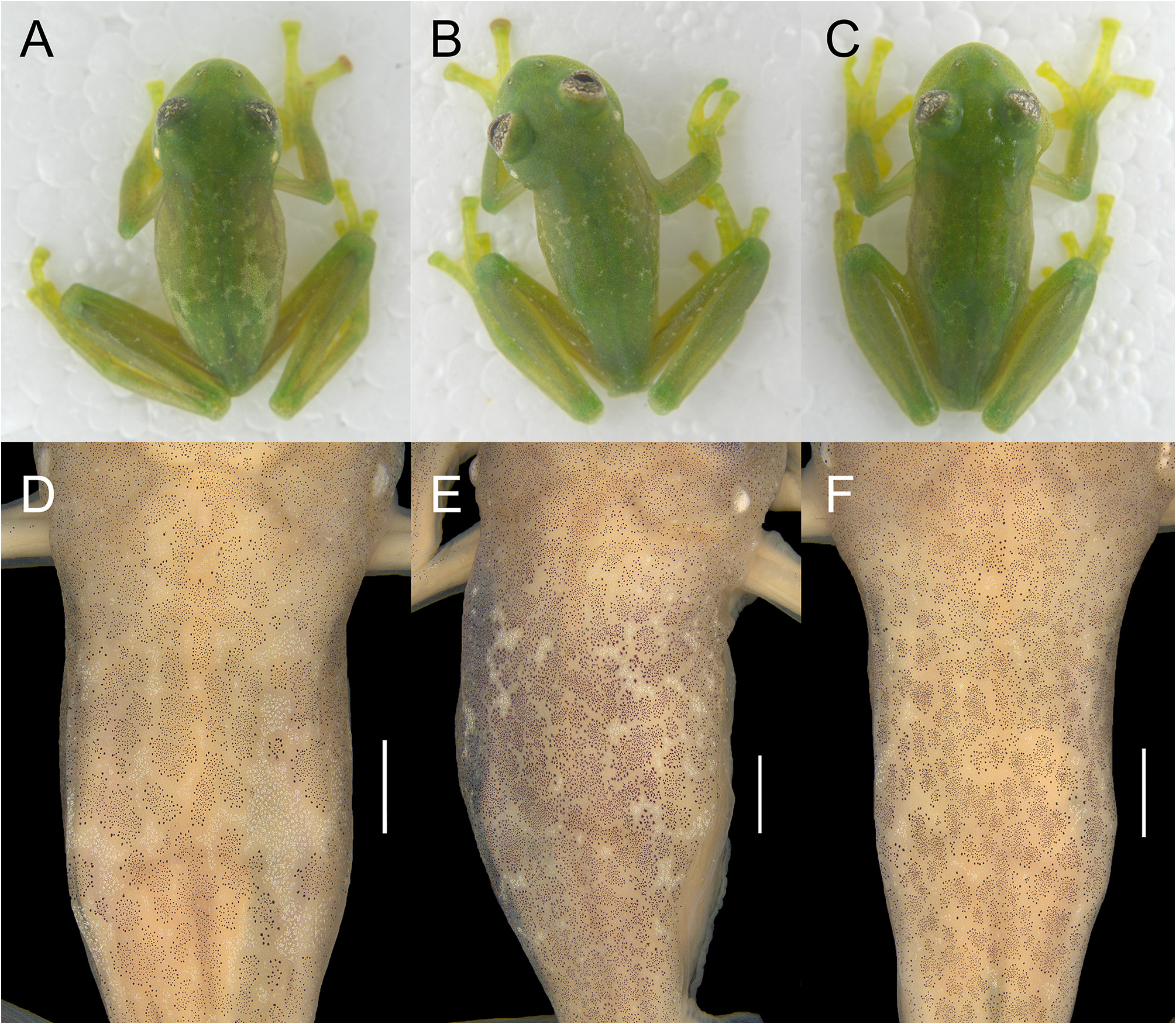
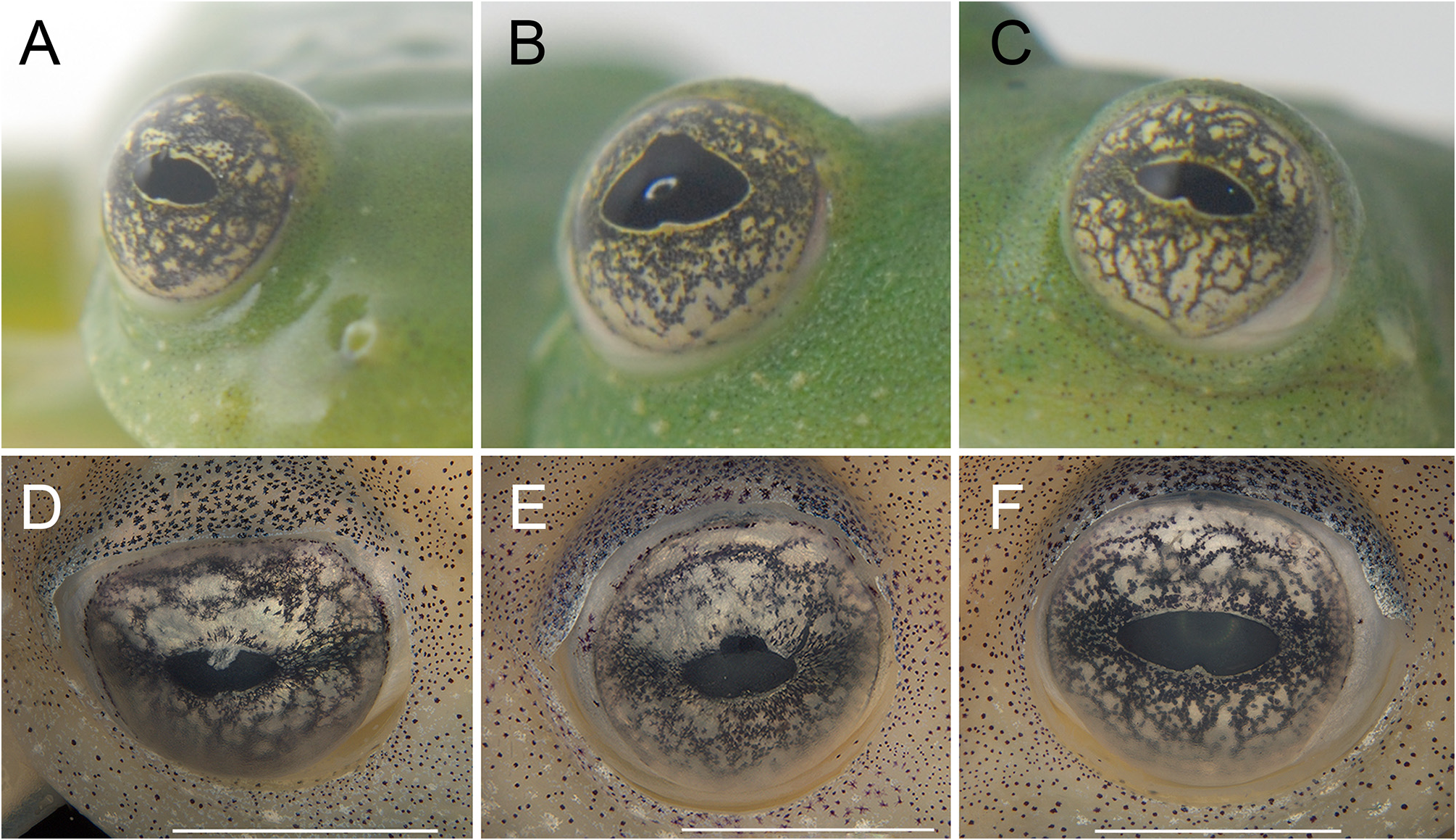
Color in life. We based this description on photographs in life of all collected topotypic individuals, except for MCP 15459. Dorsum of body, head, arms, and legs leaf green ( Fig. 3 View FIGURE 3 ). Dorsal pattern formed by clustered dark punctuations, creating different degrees of reticulation (slightly to strongly reticulated), and presence of iridophores, generally occurring in aggregations between the reticulations, but also found sparsely, especially in the head and flanks ( Fig. 4 View FIGURE 4 ). Hidden areas of thighs, hands, and feet yellowish green, some specimens having reddish hues on tip of fingers. Cloacal ornamentation and ulnar, tarsal, and tibial folds with enameled tubercles, as well as upper lip with tubercles forming a dashed white line. Lower half of tympanic membrane covered by a layer of iridophores, some specimens with membrane completely covered. Borders of nostril also covered by iridophores. Ventral skin completely translucent, allowing the visualization of internal organs. Heart, liver, and urinary bladder covered by white iridophores. Bones green. Iris background yellowish white, with dark brown reticulations, occurrence of circumpupillary ring and invagination in pupil, occurring in both surfaces, only in lower surface of pupil or absent ( Fig. 5 View FIGURE 5 ; MCP 15453 has an invagination on the lower surface and an evagination on the upper surface of the right pupil).
Color in preservative. Dorsal background surfaces of body, head, and limbs cream and decorated densely by minute and some star-shaped melanophores clustered in arrangement, forming a slightly to strongly reticulated pattern ( Fig. 4 View FIGURE 4 ). Ventral portion and flanks cream, immaculate. Iris background silvery-white with dark grey punctuations forming thin and/or thick reticulations, with concentration in the laterals of pupil ( Fig. 5 View FIGURE 5 ). Parietal peritoneum translucent, peritonea of heart, liver, and urogenital tract (kidneys dorsal surface, urinary bladder, and testes) with white iridophores; peritonea of stomach, intestines, and gallbladder translucent ( Fig. 1 View FIGURE 1 ).
Advertisement call. The advertisement call of Vitreorana parvula is pulsed, sounding like a “trii” ( Tables 2–3 View TABLE 2 View TABLE 3 ; Fig. 8 View FIGURE 8 ; see Duarte-Marin et al. 2022 Appendix 1). Calls have 1 to 5 pulses (3.2 ± 0.8 pulses), being normally single, but also grouped into 2 to 9 calls (3.8 ± 2.5 calls). Duration of grouped calls varies from 0.28 to 1.77 s (0.78 ± 0.55 s), increasing proportionally to the number of calls in the group. The silence interval between calls in the group is 165.2 to 382.3 ms (244.9 ± 52.4 ms). The call duration ranges from 4.3 to 43.2 ms (24.1 ± 6.5 ms). Call duration also increases proportionally with the number of pulses. Call repetition rate is 0.5–19.5 calls/min (11.0 ± 5.5 calls/min). Call rise is usually shorter than call fall. Call rise presents an exponential (57.3%) or logarithmic (42.7%) shape, while fall usually has a linear shape. The amplitude peak of the calls is at around the first fifth (2.8–84.7%, 22.3 ± 17.2%).
The call envelope is generally triangular pointed right (85.1%). Two calls with three pulses and one with four pulses present a variant envelope, in which the first and third pulses have similar amplitude, and the pulse in the middle of these has a lower amplitude, forming a bow tie-like shape. Calls with fairly elliptic or triangular pointed left envelopes have call fall generally shorter and amplitude peak is at around the middle of the call duration. More than 50% of the call energy is concentrated at about 43% ± 11% (12.9–73.9%) of the call duration, and in calls with one pulse, it has a slightly smaller average (ca. 30%). Calls have pulse periodic amplitude modulation (with silence intervals present between pulses). Pulses last 1.1–13.8 ms (5.6 ± 1.6 ms). Usually, pulses have an abrupt rise at 0.2–4.5 ms (1.4 ± 0.4 ms), with exponential (48.6%) or logarithmic (45.7%) shape and amplitude peak around its first fourth (7.5–83.3%, 24.9 ± 7.9%). In calls with five pulses, the last pulse has an amplitude peak around its first third. Pulse fall is generally more gradual than rise (0.6–12.3 ms; 4.3 ± 1.5 ms), but also commonly presents an exponential shape (70.3%). Pulses usually present a triangular pointed right envelope (84.7%). Symmetrical/fairly symmetrical pulses have slightly shorter pulse duration and triangular pointed left has greater pulse rise. Some pulses have a very short sustain at a maximum of 0.5 ms.
Calls normally present short intervals between pulses, equivalent to ca. 0.5 times the pulse duration (0.1–6.9 ms; 2.9 ± 1.2 ms). Grouped calls with 5 pulses have silence intervals between pulses lasting about the same as pulse duration (i.e. silence intervals ca. 1 time pulse duration). We cannot observe a clear silence interval between pulses in a few calls. Pulse repetition rate is between 88.9–232.6 pulses/s (133.0 ± 16.8 pulses/s). The period between pulse amplitude peak is 4.0–10.9 ms (8.6 ± 0.8 ms). Usually, the first or second pulse has greater amplitude. The pulse with lower amplitude is usually the last. Calls with four or five pulses have the second and first pulses with greatest and lowest amplitude, respectively. Triangular pointed left calls generally have the third and first pulses with the greatest and lowest amplitude, respectively. In calls with fairly elliptic envelope, the second pulse is normally greater. Pulses have a slight frequency modulation.
Calls have 1–4 harmonic series (3.4 ± 0.7 harmonics). The dominant frequency is always the fundamental frequency, varying from 4125–5063 Hz (4576 ± 243 Hz). Minimum and maximum frequency varies, respectively, from 3797–4922 Hz (4344 ± 213 Hz) and 4359–5297 Hz (4821 ± 224 Hz). Bandwidth varies from 281 to 750 Hz (477 ± 70 Hz). The center frequency has similar values to the dominant frequency (4125–5063 Hz; 4578 ± 228 Hz). Average entropy varies from 2.885 to 4.551 bits (3.627 ± 0.231 bits). The dominant harmonic is always the first. Second, third, and fourth harmonics have frequencies between 6656–10266 Hz (8782 ± 648 Hz), 10641–15000 Hz (13583 ± 709 Hz), and 15938–19969 Hz (18162 ± 878 Hz), respectively.
Most wave periods present subharmonics, especially between first and second harmonic, and jump frequency at the beginning and end of the call, normally in pulses with low/very low amplitude. The frequency band has a general upward frequency modulation but sometimes presents a general downward. There is no periodic frequency modulation throughout the call.
Variation. The morphological variation that we found among the topotypic specimens overlaps with what has been reported in the literature for V. uranoscopa , which is rather subtle and gradual, like a more rounded snout in dorsal view, lower folds and some larger or smaller values related to continuous acoustic or morphometric characters. Females tend to be larger than males ( Table 1 View TABLE 1 ).
Diagnosis. We compared Vitreorana parvula with all other Vitreorana species, especially those from the Atlantic Forest, based on our new description. Vitreorana parvula is distinguishable from all other Vitreorana species by its developed and crenulated outer tarsal and ulnar folds (outer tarsal and ulnar folds low or smooth); occurrence of enameled tubercles on tibia forming a fold-like structure ( Fig. 3 View FIGURE 3 ; tibia without enameled tubercles); and pair of developed horizontal flaps below and to either side of the cloaca (subcloacal ornamentation with pair of low folds or smooth).
Moreover, in Vitreorana parvula , its transparent gastrointestinal peritoneum readily distinguishes it from V. antisthenesi , V. baliomma , V. castroviejoi , V. gorzulae , V. helenae , and V. ritae (white gastrointestinal peritoneum), as well as white urinary bladder peritoneum (transparent urinary bladder peritoneum), and lower portion of tympanum with enameled iridophores in life (iridophores absent, similar coloration as surrounding skin). The completely transparent parietal peritoneum of V. parvula can also differentiate it from V. castroviejoi , V. gorzulae , V. helenae , and V. ritae (at least anterior 1/3 white).
Among the Atlantic Forest species, Vitreorana parvula differs from V. assuh by having upper lip flared (upper lip not flared); presence of microspicules, only observed by stereomicroscope (presence of spicules observed by naked eye); dentigerous process of vomer lacking teeth (presence of vomerine teeth); occurrence of enameled iridophores on dorsum (absence of enameled iridophores on dorsum); absence of flecks in dorsum (presence of flecks in dorsum); absence of melanophores on dorsal portion of Finger and Toe III (presence of melanophores in Finger and Toe III); body size small to minute, SVL <28.0 mm (body size large, SVL> 30.9). In addition, diagnostic osteological characters are presented in Zucchetti et al. (2023).
Vitreorana parvula differs from V. baliomma by having a thin white supralabial line (upper lip without supralabial line); upper lip flared (upper lip not flared); iris pattern composed by punctuations forming thin and/or thick reticulations (densely grouped lavender spots not forming reticulations); snout subovoid in dorsal and sloping in lateral view (truncated in both views); tympanic annulus and membrane clearly visible (tympanic annulus barely or non-visible); tympanic membrane at least partially covered by enameled iridophores (tympanic membrane without enameled iridophores); melanophores clustered forming reticulations (dorsal pattern with melanophores uniformly distributed); nuptial pad elongated (nuptial pad rounded); Finger II shorter than Finger III (Finger II longer or equal to Finger III); call duration ranging from 4.3–43.2 ms (call duration ranging from 185–198 ms; Bang et al. 2020); pulse emitted at a rate of 62.3–232.6 pulses/s (pulse repetition rate of 35.3–37.8 pulses/s; Bang et al. 2020).
Vitreorana parvula is distinguishable from V. eurygnatha by having the upper lip flared (upper lip not flared); snout subovoid in dorsal and sloping in lateral view (rounded or truncate in both views); tympanic annulus and membrane clearly visible (tympanic annulus normally barely visible); tympanic membrane at least partially covered by enameled iridophores (tympanic membrane without enameled iridophores); melanophores clustered forming reticulation (dorsal pattern with melanophores uniformly distributed); call duration raging from 4.3–43.2 ms (call duration ranging from 135–256 ms; Bang et al. 2020).
Vitreorana parvula differs from V. franciscana by the occurrence of enameled iridophores on dorsum (absence of enameled iridophores on dorsum); absence of pair of enlarged subcloacal warts (present); developed and crenulated outer tarsal and ulnar folds (outer tarsal and ulnar folds low); occurrence of enameled tubercles on tibia forming a fold-like (tibia without enameled tubercles); pair of developed horizontal flaps below and to either side of the cloaca (subcloacal ornamentation with pair of low folds).
In addition, Vitreorana parvula is the only species of the genus found in the southernmost Atlantic Forest (states of Rio Grande do Sul and Santa Catarina, Brazil, and Missiones province, Argentina) after more than 120 years of scientific inventories.
Natural history. In Lages, Vitreorana parvula occurs in creeks and streams inside a secondary-growth Araucaria forest, a formation commonly found in high-altitude areas of the southern Atlantic Forest. We found individuals during the night, with males starting call activity right after dusk (~19:30h). Males were found calling always in the upper side of leaves, being more active on rainy days. We observed a combat behavior between males, like the one reported by Fernandes et al. (2018). Males were found in amplexus-like position, with the top male raising the posterior part of its body and rotating while the bottom male tried to unclasp. Afterward, they grappled to the border of the leaf and finished with the bottom male climbing on the leaf and pushing the other with his legs, making him fall off. The winning male came back to its position on the leaf and resumed calling ( Fig. 11 View FIGURE 11 ).
We found four clutches in different stages of development, all deposited in the upper side of leaves, inside or on the borders of the stream bed ( Fig. 12 View FIGURE 12 ). Clutches had 25 to 27 eggs with a jelly-rich matrix surrounding them. Early-stage clutches showed eggs with general cream coloration, a light brown portion in the animal pole, and presence of empty capsules. Late-stage clutches showed developed embryos with light brown coloration and reddish hues, presenting a yellowish cream vitellus ( Fig. 12 View FIGURE 12 ). Clutches are similar to those illustrated by Machado et al. (2014). We only found Rhinella icterica occurring in syntopy with V. parvula in one of the sampled streams. More information on the reproductive behavior (courtship, amplexus, oviposion, and maternal care) of V. parvula remains unknown.
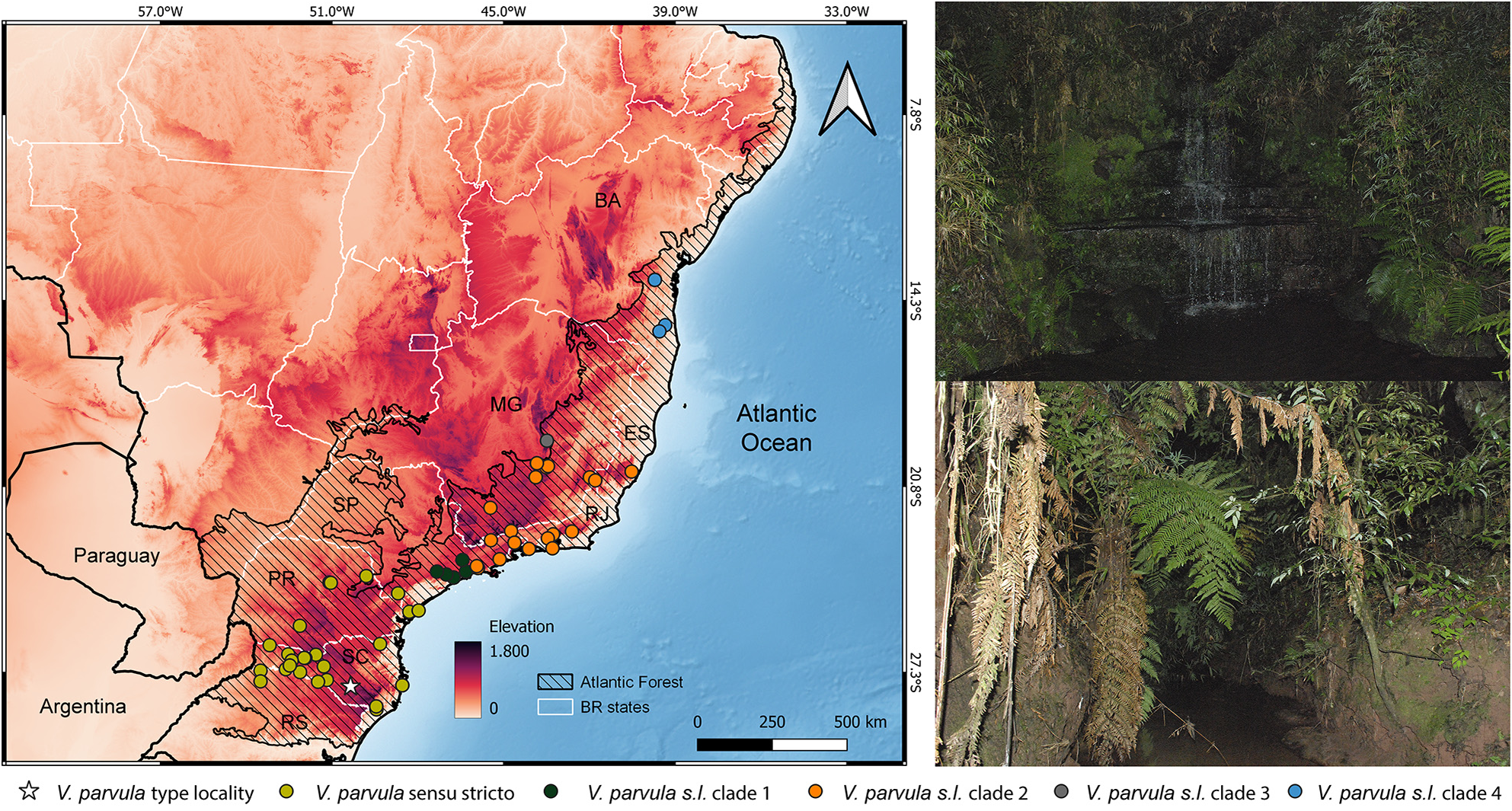
Geographic distribution. The distribution of Vitreorana parvula encompasses the distribution previously defined for V. uranoscopa , occupying forested streams, creeks, and waterfalls throughout the Atlantic Forest extension, from the south of Bahia to the north of Rio Grande do Sul and part of Argentina, from about sea level to high altitudinal areas (≥ 1800 m a.s.l.; Fig. 13 View FIGURE 13 ). It is likely to occur in Paraguay on the border with Argentina and Brazil ( Brusquetti & Lavilla 2006).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Vitreorana parvula ( Boulenger, 1895 )
| Zucchetti, Victor M. & Castroviejo-Fisher, Santiago 2024 |
Vitreorana uranoscopa
| Guayasamin, J. M. & Castroviejo-Fisher, S. & Trueb, L. & Rada, M. & Vila, C. 2009: 39 |
Hyalinobatrachium parvulum
| Ruiz-Carranza, P. M. & Lynch, J. D. 1991: 24 |
Hyalinobatrachium uranoscopum
| Ruiz-Carranza, P. M. & Lynch, J. D. 1991: 25 |
Centrolenella albotunica
| Duellman, W. E. 1977: 63 |
Centrolenella dubia
| Duellman, W. E. 1977: 188 |
Centrolenella lutzorum
| Duellman, W. E. 1977: 191 |
Centrolenella parvula
| Duellman, W. E. 1977: 192 |
Centrolenella uranoscopa
| Duellman, W. E. 1977: 196 |
Centrolenella vanzolinii
| Duellman, W. E. 1977: 196 |
Cochranella albotunica
| Taylor, E. H. & Cochran, D. M. 1953: 1648 |
Cochranella dubia
| Taylor, E. H. & Cochran, D. M. 1953: 1653 |
Cochranella lutzorum
| Taylor, E. H. & Cochran, D. M. 1953: 1638 |
Cochranella parvula
| Taylor, E. H. & Cochran, D. M. 1953: 1650 |
Cochranella uranoscopa
| Taylor, E. H. & Cochran, D. M. 1953: 1635 |
Cochranella vanzolinii
| Taylor, E. H. & Cochran, D. M. 1953: 1646 |
Centrolenella uranoscopa
| Noble, G. K. 1926: 18 |
Hylella parvula
| Boulenger, G. A. 1895: 646 |