
Quedius mutilatus Eppelsheim, 1888
|
publication ID |
https://doi.org/10.5852/ejt.2018.401 |
|
publication LSID |
lsid:zoobank.org:pub:9C9DB157-AAA5-40B7-BA0B-9A57779382C1 |
|
DOI |
https://doi.org/10.5281/zenodo.5695864 |
|
persistent identifier |
https://treatment.plazi.org/id/03D5FE7E-7048-BF26-FDCD-FD70FD45FA17 |
|
treatment provided by |
Plazi |
|
scientific name |
Quedius mutilatus Eppelsheim, 1888 |
| status |
|
Quedius mutilatus Eppelsheim, 1888 View in CoL
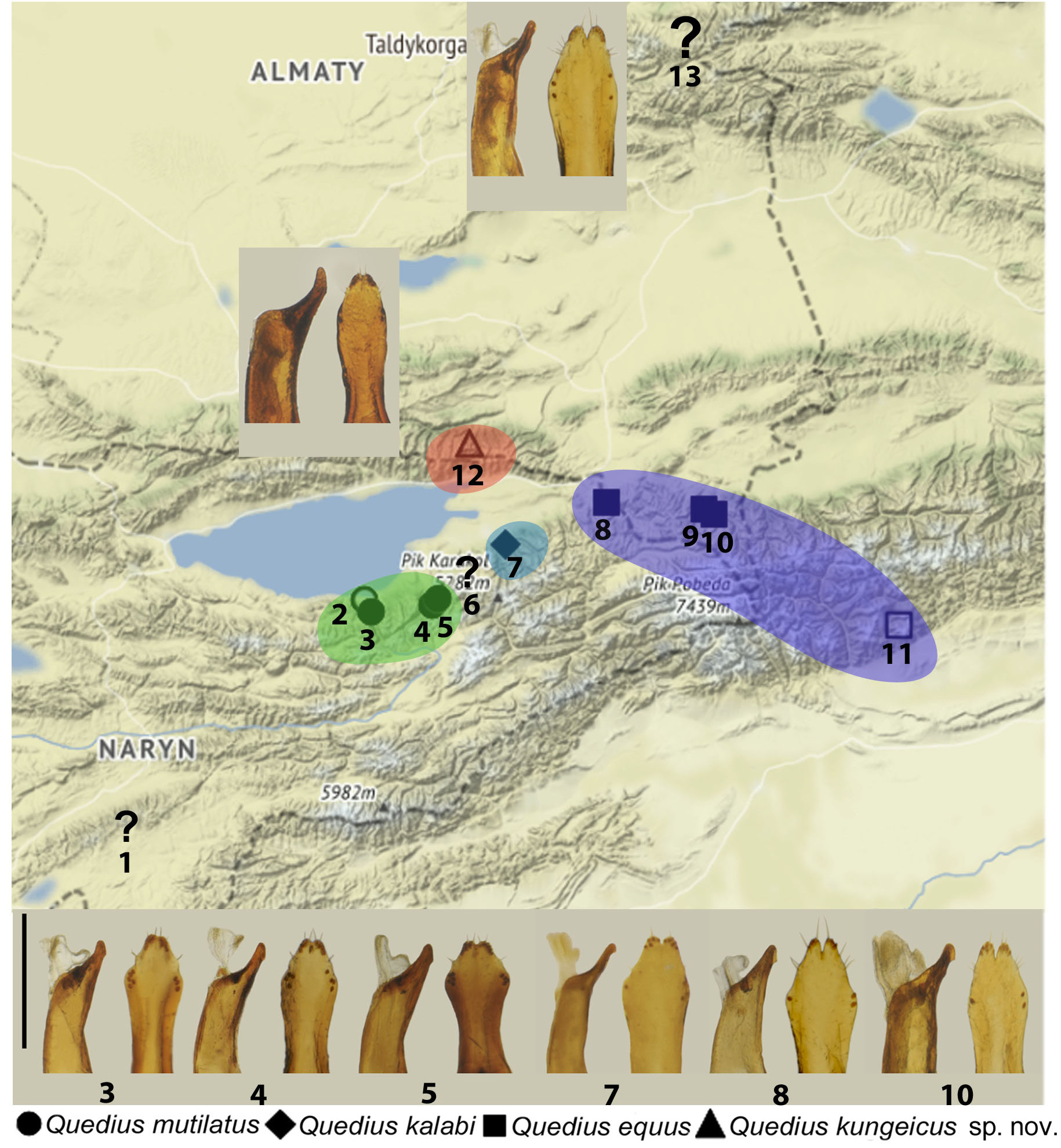
Figs 1–2 View Fig.1 View Fig. 2 , 4A–B View Fig. 4
Quedius mutilatus Eppelsheim, 1888: 58 View in CoL (original description).
Quedius mutilatus View in CoL – Gridelli 1924: 23 (redescription). — Coiffait 1978: 161 (redescription). — Smetana 1998: 118 ( lectotype designation, illustrations, comparison with Q. kalabi View in CoL ).
Material examined
Type material
KYRGYZSTAN: Terskei Alatau mountain range , lectotype, ♂, “ ♂ / 245 [handwritten label]/ Fluss. Tamgi [river Tamga in Karakol Gorge] Turkestan. Leg. Akinin [handwritten label]/ c.Epplsh. Steind. d [printed label]/ Q. mutilatus Epp. Type [handwritten label] /TYPUS [red printed label]/ LECTOTYPE Quedius mutilatus Eppelsheim A. Smetana des. 1997 [red handwritten label]” ( NMW).
Additional material
KYRGYZSTAN: Terskei Alatau mountain range : 2 ♂♂, Barskoon Valley [ 42.0900° N, 77.5900° E], 14–15 Jul. 1983, S. Alekseev leg. ( cRyv); GoogleMaps 4 ♂♂, 4 ♀♀, same collecting data as preceding, but 2400 m, 18 Jun. 1997, V. Dolin leg. ( NMW, ZMUC); GoogleMaps 2 ♂♂, Chon-Kyzyl-Suu River , 3000 m [ 42.1700° N, 78.2500° E], 29 Jul. 1955, D. Panfilov leg. ( ZIN); GoogleMaps 4 ♂♂, same collecting data as preceding, but 2500– 2700 m, coniferous forest, 2 Jul. 1959, Zaslavsky leg. ( ZMUC); GoogleMaps 2 ♂♂, 3 ♀♀, at Kochevnikov stationary ( 25 km S of village Pokrovka [village Kyzyl-Suu]), 2450 m [ 42.1260° N, 78.0250° E], coniferous forest, leaf litter under moss and grass, 2–7 Jul. 1988, V. Yanushev leg. ( cRyv); GoogleMaps 4 ♂♂, 2 ♀♀, same collecting data as preceding, but 27–28 Jun. 1988 ( cRyv); GoogleMaps 2 ♂♂, 4 ♀♀, same collecting data as preceding, but coniferous forest, 2500 m, 12–31 Aug. 1984, N. Turtseva leg. ( cRyv, ZMUC); GoogleMaps 1 ♂, 4 ♀♀, same collecting data as preceding, but 15–18 Jun. 1985 ( cRyv); GoogleMaps 2 ♀♀, same collecting data as preceding, but 19 Jun. 1986 ( cRyv); GoogleMaps 4 ♂♂, ‘ Turkestan Akinin-Tepe’ ( NMW).
Redescription
MEASUREMENTS AND RATIOS. Lectotype: HL: 1.07, HW: 1.11, PL: 1.30, PW: 1.52, EL: 1.16, EW: 1.57, FB: 3.52, HL/ HW: 0.96, PL/ PW: 0.85, EL/EW: 0.74; non-type material (range, arithmetic mean, n = 30): HL: 1.18–1.64 (1.36), HW: 1.11–1.57 (1.29), PL: 1.32–.82 (1.50), PW: 1.48–2.02 (1.74), EL: 1.25– 1.70 (1.42), EW: 1.52–2.00 (1.70), FB: 3.77–5.07 (4.28), HL/HW: 0.93–1.22 (1.03), PL/PW: 0.79–0.95 (0.87), EL/EW: 0.75–0.93 (0.83).
BODY. Length 8–12 mm; brown to light brown including appendages, elytra and abdominal tergites, becoming slightly paler toward apex, sometimes head and abdomen slightly darker than pronotum and elytra; distinctly flattened dorso-ventrally.
HEAD. About as long as wide [HL/HW: 0.93–1.22 (1.03)], with broadly arcuate posterior angles; surface with distinct microsculpture of transverse waves on disc, becoming more or less isodiametric on frons between eyes, and with distinct sparse non-setiferous micropunctation. Eyes small and flat, not protruding over lateral contour of head; temples about 2.00–3.18 (2.54) times as long as longitudinal diameter of eye. Head disc on each side with setiferous punctures as following: anterior frontal puncture at inner margin of eye, posterior frontal puncture very close to neck, and pair of smaller vertical punctures immediately medio-posteriorly from the latter puncture. Each temple with two temporal punctures of which posterior temporal puncture closer to posterior margin of head than to posterior margin of eye; additionally, temples with some fine setiferous punctures bearing short pale setae.
ANTENNAE. Segments as following: second and third combined as long as first; third slightly longer than second; fourth and fifth, each, as long as wide; sixth to tenth, each, wider than long and gradually widening towards apex of antenna; last segment slightly shorter than two preceding segments combined.
PRONOTUM. Wider than long [PL/PW: 0.79– 0.95 (0.86)] widest at about its middle, gradually narrowing anteriad and vaguely posteriad; anterior lateral portions sloping ventrad, posterior lateral portions slightly explanate; both anterior and posterior pronotal angles distinct; disc with microsculpture and non-setiferous micropunctation similar to that on posterior part of head. Disc with three setiferous punctures in each dorsal row: one puncture before middle of pronotum, one close to its anterior margin, and one (smaller) latero-anteriorly at pronotal anterior margin; sometimes some punctures weak or absent, in both or one row, or sometimes smaller additional puncture near basalmost discal puncture. Antero-lateral area of pronotal disc with one to two sublateral punctures situated slightly anterior to large lateral puncture.
SCUTELLUM. Impunctate, with faint transverse microsculpture.
ELYTRA. Parallel-sided, relatively short [EL/EW: 0.74–1.12 (0.83)], about as long as pronotum [PL/EL: 0.98–1.20 (1.06)], and slightly narrower than pronotum at widest point [PW/EW: 0.94–1.34 (1.03)]; setiferous punctation fine and moderately dense with interspaces larger than punctures diameter, shiny, with distinct minute irregularities; pubescence yellowish to brownish. Wings vestigial.
ABDOMEN. With fine and dense punctation; interspaces with minute irregularities; posterior margin of tergite VII without palisade fringe ( Fig. 2A View Fig. 2 ).
Male
SECONDARY SEXUAL CHARACTERS. First three segments of front tarsus more strongly dilated than in females; second segment slightly wider or as wide as apex of tibia. Sternite VIII with distinct roughly triangular medio-apical emargination; tergite X triangular, strongly narrowed apically with few strong and numerous smaller apical setae; sternite IX moderately elongate with wide and glabrous basal, and sparsely setose and vaguely bilobed apical portion.
AEDEAGUS ( Fig. 2B View Fig. 2 ). Median lobe: (in parameral view) parallel-sided, with obtusely pointed apex; (in parameral or lateral view) with minute tooth situated near its apex. Paramere (in parameral view) as wide as median lobe, lanceolate, with slightly incised apex; underside with irregular groups of sensory peg setae, including a pair of apical groups with ca 3–7 peg setae in each group, and a pair of lateral groups with ca 4–6 peg setae each; additionally, parameral apex with normal setae including pair of apical setae on each side of apical incision and ca 1–4 lateral setae on each side below. Internal sac without large, strongly sclerotized structures.
Female
SECONDARY SEXUAL CHARACTERS. First four segments of protarsi dilated, but slightly narrower than in males. Tergite X triangular, gradually narrowed apically, pigmented and with long setae medioapically.
Distribution and bionomics
Quedius mutilatus is endemic to the central Terskei Alatau mountain range, where it is known from Karakol Gorge in the west ( type locality) to the Chon-Kyzyl-Suu River in the east. Examined material has been collected from the second half of May through the end of August, all at high elevations around 2000–3000 m. At least some specimens without bionomic records were presumably found under stones as a by-catch by various entomologists, including specialists targeting Carabidae and Elateridae who, as a rule, intensively turn stones at elevations above the timber line. A few specimens with recorded bionomics have been found in coniferous leaf litter and other ground-based debris like grass or moss.
Some specimens from NMW bear clearly written, but ambiguous geographical labels with ‘Turkestan Akinin-Tepe’, which we could not interpret in spite of consulting historical literature about I.Y. Akinin’s travels in Middle Asia ( Jacobson 1902). Based on the structure of the aedeagus, though, these specimens clearly belong to Q. mutilatus .
Comparison
Quedius mutilatus differs from Q. kungeicus sp. nov. by the rhomboid (as opposed to ovoid) apical portion of the paramere and a slight, but distinct incision on its apex (as opposed to indistinct incision; aedeagus in parameral view), as well as the less curved apical portion of the median lobe, with stronger ventral sub-apical tooth (aedeagus in lateral view). From Q. kalabi it differs by the more robust and less curved apical portion of the median lobe with its shorter apical part (aedeagus in lateral view) and, usually, by the larger number of sensory peg setae in lateral groups on the paramere. From Q. equus it differs by the less incised apex of the paramere and a distinctly greater number of sensory peg setae in the lateral groups on the paramere.
Notes on the type material
Quedius mutilatus Eppelsheim, 1888 was described based on two specimens with unclearly recorded collecting localities. Of these, a male comes from ‘Fluss. Tamgi’ which we interpret as River Tamga in the Karakol Gorge of Terskei Alatau, while a female comes from ‘Lake Issyk-Kul’, an even less precisely outlined area around a large lake in Kyrgyzstan. Smetana (1998), who designated the male as a lectotype, interpreted the female paralectotype label literally as the Lake Issyk-Kul at ca 1500 m of elevation and suggested that, unlike the high altitudinal Q. kalabi , Q. mutilatus is confined to lower elevations. However, such an interpretation is not confirmed here with new and better georeferenced material.
Before Smetana’s lectotype designation and his latest taxonomic treatment of this species, the male syntype was examined by Gridelli (1924), who noted some morphological characters, and by Coiffait (1978), who first illustrated the aedeagus for Q. mutilatus .
It is noteworthy that the paralectotype female of Q. mutilatus from ‘See Isyk-Kul [Lake Issyk-Kul]’ is teneral, slightly smaller than the lectotype and notably smaller than the size range of the non-type specimens. Given its small size and a lack of precise locality information, the species identity of the paralectotype of Q. mutilatus remains unclear. Therefore, we list it among other undetermined material of the Q. mutilatus group at the end of this paper.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Staphylininae |
|
Tribe |
Staphylinini |
|
SubTribe |
Quediina |
|
Genus |
|
|
SubGenus |
Microsaurus |
Quedius mutilatus Eppelsheim, 1888
| Salnitska, Maria & Solodovnikov, Alexey 2018 |
Quedius mutilatus
| Smetana 1998: 118 |
| Coiffait 1978: 161 |
| Gridelli 1924: 23 |
Quedius mutilatus Eppelsheim, 1888 : 58
| Eppelsheim 1888: 58 |