
Uperodon rohani, Garg & Senevirathne & Wijayathilaka & Phuge & Deuti & Manamendra-Arachchi & Meegaskumbura & Biju, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4384.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:18DD1320-7914-4E09-A46C-707069DC69F5 |
|
DOI |
https://doi.org/10.5281/zenodo.5587723 |
|
persistent identifier |
https://treatment.plazi.org/id/D612A25A-E09E-4193-8B89-3DC62746FB55 |
|
taxon LSID |
lsid:zoobank.org:act:D612A25A-E09E-4193-8B89-3DC62746FB55 |
|
treatment provided by |
Plazi |
|
scientific name |
Uperodon rohani |
| status |
sp. nov. |
Uperodon rohani View in CoL sp. nov.
Rohan’s Globular Frog
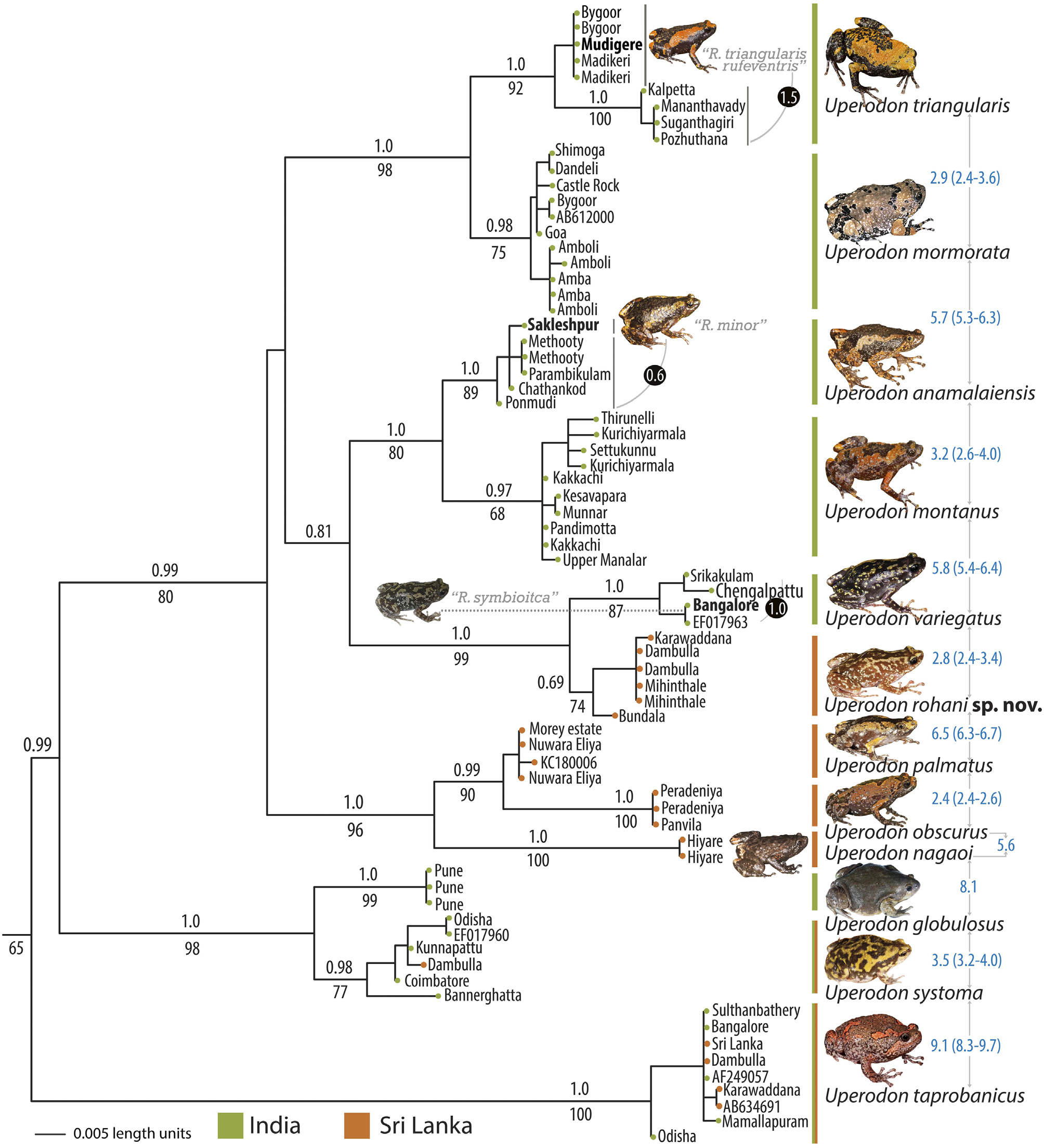
( Figs. 1 View FIGURE 1 , 2A, 2D View FIGURE 2 , 3L View FIGURE 3 , 5K View FIGURE 5 , 12 View FIGURE12 , 13 View FIGURE 13 , 14 View FIGURE14 ; Tables 1–4) http://zoobank.org/urn:lsid:zoobank.org:act:D612A25A-E09E-4193-8B89-3DC62746FB55
Holotype. DZ 1449, an adult male, from Mihintale, Anuradhapura district , Sri Lanka, collected by Madhava Meegaskumbura and team, on 0 2 October 2014 . Paratypes. DZ 1448, DZ 1450 and DZ 1374, three adult males, collected along with the holotype; DZ 1336, an adult male, from Bundala NP, Hambantota district , Sri Lanka, collected by MM and team, on 0 4 November 2013 ; DZ 1520 and DZ 1521, two adult males, and DZ 1425, an adult female, from Makandura, Kurunegala district , Sri Lanka, collected by MM and team, on 11 September 2014 ; DZ 1486, an adult male, and DZ 1485, an adult female, from Ampara, Ampara district , Sri Lanka, collected by MM and team, on 10 October 2012 ; DZ 1129–DZ 1132, four adult females, from Dambulla, Matale district , Sri Lanka, collected by MM and team, on 0 4 January 2013 . Referred specimens. NHM 1931.2 About NHM .1.21, an adult male, from “ Ceylon ”, Sri Lanka, collected by GH Herry ; NHM 1973.3015 About NHM , an adult male, from Kurunegala, “ Kurunegala dist. ,” Sri Lanka, collected by R Sananayake; NMH 1976.264 – NHM 1976.265 About NHM , two adult males, from Badulla, “ Hambantota dist.,” Sri Lanka, collected by JC Naggs; WHT 5320, an adult male, from Nochchiyagama , Anuradhapura district , Sri Lanka, collected by MM Bahir, C Gans and C Yahalmulla, on 18 November 2000 ; WHT 0 1188, an adult male, from Sigiriya, Near Rest house, Matale district , Sri Lanka, collected by D Gabadge, on 0 8 February 1996 ; WHT 1749, an adult male, from Polonnaruwa, Polonnaruwa district , Sri Lanka, collected by A Nollert, on 28 July 1997 ; WHT 0 1189, an adult male, from Anuradhapura, Anuradhapura district , Sri Lanka, collected by D Gabadage and MM Bahir, on 11 August 1995 ; NHM 1973.3014 About NHM , an adult female, from Kurunegala: Ceylon, “ Kurunegala dist. ,” Sri Lanka, collected by R Sananayake; WHT 0 1191, an adult female, from Palatupana, Kirinda, Hambantota district, Sri Lanka, collected by U. Premachandra, in August 1995 .
Etymology. This species is named after Rohan Pethiyagoda, in recognition of his contribution to biodiversity studies in India and Sri Lanka, especially on frogs and fishes. The species epithet is formed as a noun in the genitive case.
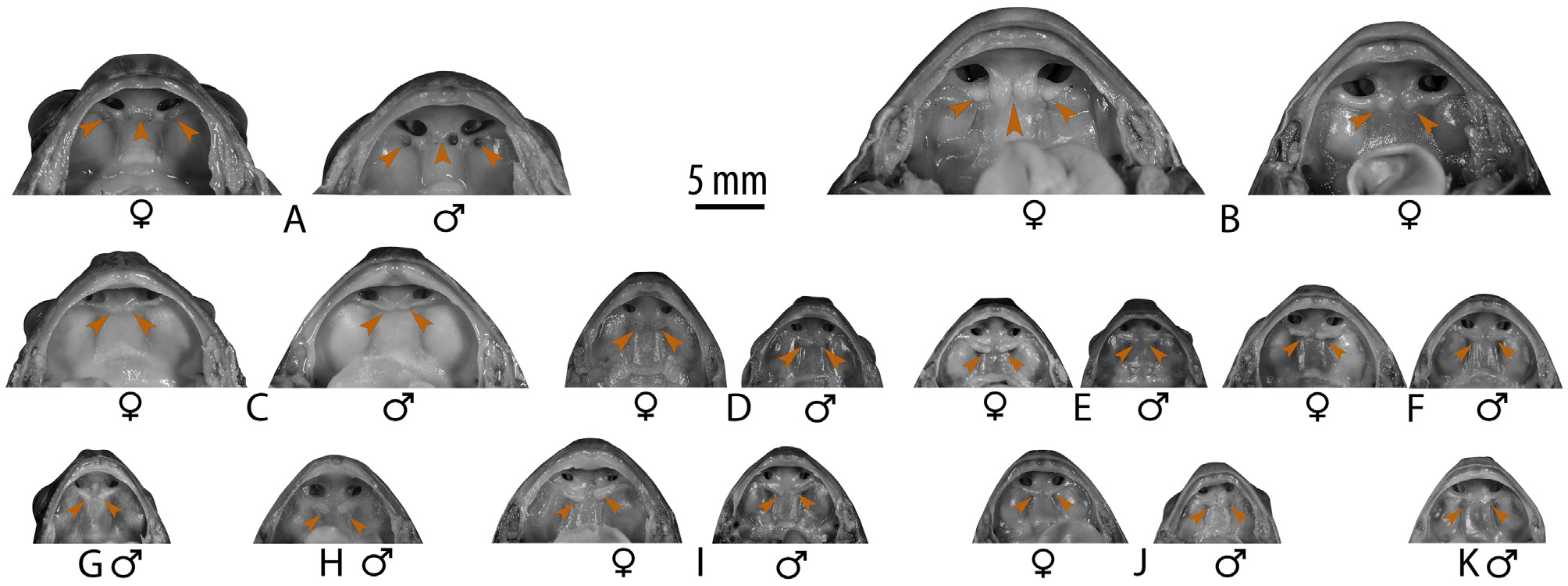
Comparison. Uperodon rohani sp. nov. cannot be confused with U. globulosus , U. systoma and U. taprobanicus due to its smaller snout-vent size, male SVL 26–34 mm, N = 16, female SVL 28–35 mm, N = 8 (vs. larger, U. globulosus male SVL 51–56 mm, N = 2, female SVL 66–72 mm, N = 3; U. systoma male SVL 51–56 mm, N = 2, female SVL 47–64 mm, N = 4; and U. taprobanicus male SVL 42–52 mm, N = 3, female SVL 53–55 mm, N = 2). Uperodon rohani sp. nov. specifically also differs from the former two by its finger tips having truncate discs (vs. rounded without discs), and presence of two weakly-developed neopalatinal ridges on posterior side of each choana, fused with the vomerine odontophores (vs. two prominent dermal projections on the neopalatinal ridges on posterior side of each choana, separated from the vomerine odontophores); and from U. taprobanicus by presence of two weakly-developed neopalatinal ridges on posterior side of each choana, fused with the vomerine odontophores (vs. well-developed neopalatinal ridges). Further, U. rohani sp. nov. differs from U. anamalaiensis , U. montanus , U. obscurus and U. palmatus by its ventral skin being white to off-white in life (that turns greyish-white in preservation) without any spots or blotches (vs. light or dark brown with prominent spots or blotches), and absence of webbing between toes in both the sexes (vs. present). Uperodon rohani sp. nov. differs from U. nagaoi by its ventral skin being white to off-white in life (that turns greyish-white in preservation) without any spots or speckles (vs. brown to dark brown in life that turns brown or dark greyish-brown in preservation, with or without inconspicuous spots or speckles). For detailed comparison of U. rohani sp. nov. with U. globulosus , U. systoma , U. taprobanicus , U. anamalaiensis , U. montanus , U. mormorata , U. nagaoi , U. obscurus , U. palmatus and U.
triangularis see ‘comparison’ section of those species. Uperodon rohani sp. nov. is morphologically and genetically close to U. variegatus , from which it differs by its snout nearly equal to eye diameter, male SL/EL ratio 1.0–1.2, N = 16, female SL/EL ratio 0.8–1.0, N = 8 (vs. snout relatively longer than eye, male SL/EL ratio 1.2–1.6, N = 12, female SL/EL ratio 1.2–1.4, N = 7); head relatively narrower, male HW/HL 1.0–1.1, N = 16, female HW/HL 1.0–1.2, N = 8 (vs. head relatively wider, male HW/HL 1.1–1.4, N = 12, female HW/HL 1.1–1.4, N = 7); and shank relatively longer than thigh, male SHL/TL ratio 1.0–1.1, N = 8, female SHL/TL ratio 1.1, N = 8 (vs. relatively shorter, male SHL/TL ratio 0.9–1.0, N = 12, female SHL/TL ratio 0.8–0.9, N = 7) ( Table 4 View TABLE 4 ).
The new species cannot be confused with previously available names, Engystoma marmoratum Cuvier, 1829 and Systoma leschenaultii Tschudi, 1838, currently under the synonymy of Uperodon systoma Schneider, 1799 . The former was described from “l’Inde” (= India) and as stated by Peters (1864 “1863”)—“bereits richtig erkannt batten = Uperodon marmoratum D. B. und miifste daher den Species namen “ systoma ” behalten”, was already correctly recognized as Uperodon marmoratum (= Uperodon systoma ) by Duméril and Bibron (1841). The latter, also stated by Duméril and Bibron (1841) —“peut-être de la même espèce que notre Upérodonte marbré”, is perhaps the same as Uperodon systoma , from which the new species cannot be confused based on several morphological characters such as smaller body size (vs. larger), finger tips with truncate discs (vs. rounded without discs) and vomerine odontophores fused with neopalatinal ridges (vs. separated). Furthermore, three other names— Hylaedactylus carnaticus Jerdon, 1854 “1853”, Callula olivacea Günther, 1876 “1875” and Ramanella symbioitca Rao and Ramanna, 1925 , currently under the synonymy of Uperodon variegatus Stoliczka, 1872 , were also described from regions in India (“Carnatic”, “Yellagherry hills” and “Ellore”, respectively), while the distribution of U. rohani sp. nov. is restricted to Sri Lanka. For specific differences with U. systoma and U. variegatus , see the detailed comparison with those species.
Morphometric differentiation. Uperodon rohani sp. nov. showed morphological differentiation from its closely related species, U. variegatus . Principal Component Analysis (PCA) for the adult male specimens ( U. rohani sp. nov.: N = 12; U. variegatus : N = 16) recovered five Principal Components (PC) with eigenvalues>1.0 that accounted for 77.2% of the total variance; PC 1 explained 32.3% of the variations, with highest loadings for SVL, HW, IUE, IFE, MN, HAL and TL; PC 2 explained 22.1% of the variations, with highest loadings for SL, IBE, EN, SHL and FOL; PC 3 explained 10% of the variations, with highest loadings for UEW and NS; PC 4 explained 6.9% of the variations, with highest loadings for EL; PC 5 explained 6% of the variations; and the remaining factors altogether explained 22.8% of the variations ( Table S2). Principal Component Analysis (PCA) for the adult female specimens ( U. rohani sp. nov.: N = 7; U. variegatus : N = 8) recovered three Principal Components (PC) with eigenvalues>1.0 that accounted for 82.3% of the total variance; PC 1 explained 46.2% of the variations, with highest loadings for SVL, HW, SL, IUE, UEW, IFE, IBE, MN, EN and TL; PC 2 explained 26.9% of the variations, with highest loadings for HL, FAL, HAL and SHL; PC3 explained 9.2% of the variations, with highest loadings for EN; and the remaining factors altogether explained 17.7% of the variations ( Table S2). Projections of factor planes 1 and 2 showed distinct clusters for the two species ( Fig. 13 View FIGURE 13 ).
Osteological comparison. Uperodon rohani sp. nov. and its closely related species, U. variegatus , have the following skeletal features: Anteriorly slender, basally wider crania, with widest at anterior margins of tympanic annulus in both species. Cranium length is 93% of its width in U. rohani sp. nov. and 84.2% in U. variegatus . Frontoparietals are largest, covering more than half of crania, slender and quadrangular shaped in both species. Nasals, lying dorsal to nasal capsules, triangular-shaped, medial edge 20.6% of the mid-cranial width in U. variegatus and 20.9% in U. rohani sp. nov. Posterolateral margins of frontoparietals slightly overlay anterolateral margins of prootics. Major differences between the two species concentrate mainly on three bones of the lower and upper jaws—premaxillae, mentomeckelians and dentaries. Premaxillae usually composed of alary, dentary and maxillary processes. In both species, premaxillae are edentate. However, shape of dentary process differs immensely, wherein U. rohani sp. nov. has well-bifurcated dentary process with 74.9% height compared to width of premaxillae. Comparatively, dentary process of U. variegatus is rectangular in shape, with a slight median depression, and 27.4% height compared to premaxillary width. Mentomeckelians are observed as cylindrical endochondral elements, anterior to the anterior ends of Meckel’s cartilages, only in U. variegatus . Mentomeckelians are absent in U. rohani sp. nov.; however, the infrarostrals, which are the precursors of Mentomeckelians are still present and can be seen clearly ( Fig. 14 View FIGURE14 ). Development of dentaries also shows differences between the two species. Dentaries are well-developed, extend for about one-third and two-third of the length of the mandible arc in U. rohani sp. nov. and U. variegatus , respectively ( Fig. 14 View FIGURE14 ).
Genetic divergence. For 16S mitochondrial gene sequences, the sampled populations of Uperodon rohani sp. nov. showed an average intraspecific distance of 0.5% (range 0–1.4%, N = 6). The sample from Bundala showed a divergence of up to 1.4% from other populations. Genetically, U. rohani sp. nov. is closely related to U. variegatus , from which it was found to differ by an average uncorrected genetic distance of 2.8% (range 2.4–3.4%, N = 24). For interspecific genetic distances with all other members of the genus, see Table 3.
Description of holotype (measurements in mm) (3L, 5K, 12B–J). Small-sized (SVL 26.0), relatively slender adult male; head small (HW 6.7, HL 6.2, IFE 3.5, IBE 5.5), less than one-fourth (23.4%) of body length, wider than long (HW/HL ratio 1.1); snout subovoid in dorsal, subovoid to nearly truncate in ventral view, rounded in lateral view, its length (SL 2.6) slightly longer than horizontal diameter of eye (EL 2.5); loreal region rounded with vertical canthus rostralis; interorbital space wider (IUE 2.9) than upper eyelid (UEW 1.6) and internarial distance (IN 1.9); nostril closer to tip of snout (NS 0.7) than to eye (EN 1.4); supratympanic fold distinct, extending from posterior corner of upper eyelid near to insertion of forelimb at axilla; eye diameter (EL 2.5); vomerine odontophores present on the palate, without teeth; presence of two weakly-developed neopalatinal ridges on posterior side of each choana, fused with the vomerine odontophores; tongue emarginate. Forelimbs moderately long and thin; forearm (FAL 5.2) shorter than hand length (HAL 7.2); finger length formula I<II<IV<III, tips of all fingers with oval discs, discs moderately wide compared to finger width (FD I 0.9, FW I 0.4; FD II 1.3, FW II 0.4; FD III 1.2, FW III 0.4; FD IV 1.2, FW IV 0.5); subarticular tubercles prominent, oval, all present; two well-developed palmar tubercles, oval. Hind limbs relatively long and thin, thigh length (TL 9.2) slightly shorter than shank (SHL 9.4) and shorter than foot (FOL 11.1); relative digit lengths I<II<V<III<IV; tips of all toes slightly enlarged with truncate to rounded discs, discs rather wide compared to toe width (TD I 0.7, TW I 0.6; TD II 0.8, TW II 0.6; TD III 0.7, TW III 0.5; TD IV 0.7, TW IV 0.5; TD V 0.5, TW V 0.5); foot webbing absent; subarticular tubercles prominent, oval; two smooth metatarsal tubercles, oval, outer one slightly longer than the inner.
Skin of dorsum, snout, and between eyes, shagreened; sides of head, posterior part of back, and upper and lower parts of flank, smooth; dorsal surfaces of forelimb, thigh, tibia and tarsus, smooth to shagreened; ventral surfaces smooth.
Colouration. In preservation: Dorsum dark brown with irregular olive grey coloured blotches, pair of faint and irregular olive grey longitudinal streaks extending from upper eyelids up to vent; dorsal surfaces of forelimbs and hind limbs lighter in colour compared to dorsum and with faint yellowish-grey spots or blotches; sides of head brown; ventral surfaces (including limbs) greyish-white, throat with blackish-brown calling patch ( Fig. 3L View FIGURE 3 ). Colour in life: Dorsum reddish brown with pair of irregular olive yellow coloured longitudinal streaks extending from upper eyelids up to vent, scattered olive yellow coloured spots and blotches especially towards dorsolateral surfaces and posterior part of back, larger olive yellow blotch in center of dorsum, few scattered spots on head, olive yellow patch between nostrils and upper eyelids; lateral sides of head brown with scattered olive yellow spots near margins of mouth; anterior and posterior parts of groin light brown; dorsal surfaces of limbs brown to light brown with scattered olive yellow spots; throat light brown with bluish-black calling patch and irregular light grey speckles; chest, belly and thighs, greyish-white with light brown margins; limbs light brown with irregular light grey speckles ( Figs. 12F–J View FIGURE12 ).
Variations. Morphometric measurements for 16 males and eight females, including the type series, are given in Table 4 View TABLE 4 . DZ 1374 and DZ 1336: dorsal skin smooth; DZ 1129: lateral surfaces slightly granular; DZ 1374: lateral surfaces of head slightly granular. DZ 1374: dorsum coffee brown in colour with prominent irregular olive grey spots or patches; DZ 1521: dorsum dark brown with olive grey blotches; DZ 1425: dorsum dark brown with small olive grey spots scattered throughout, scattered spots also seen on surfaces of hand, foot, sides of head and groin; DZ 1485: dorsum darker brown with fewer blotches.
Secondary sexual characters. Male (DZ 1449): vocal sac externally visible on the lower jaw; female (DZ 1425): ova white, pigmented on poles (diameter 0.6–1.0 mm).
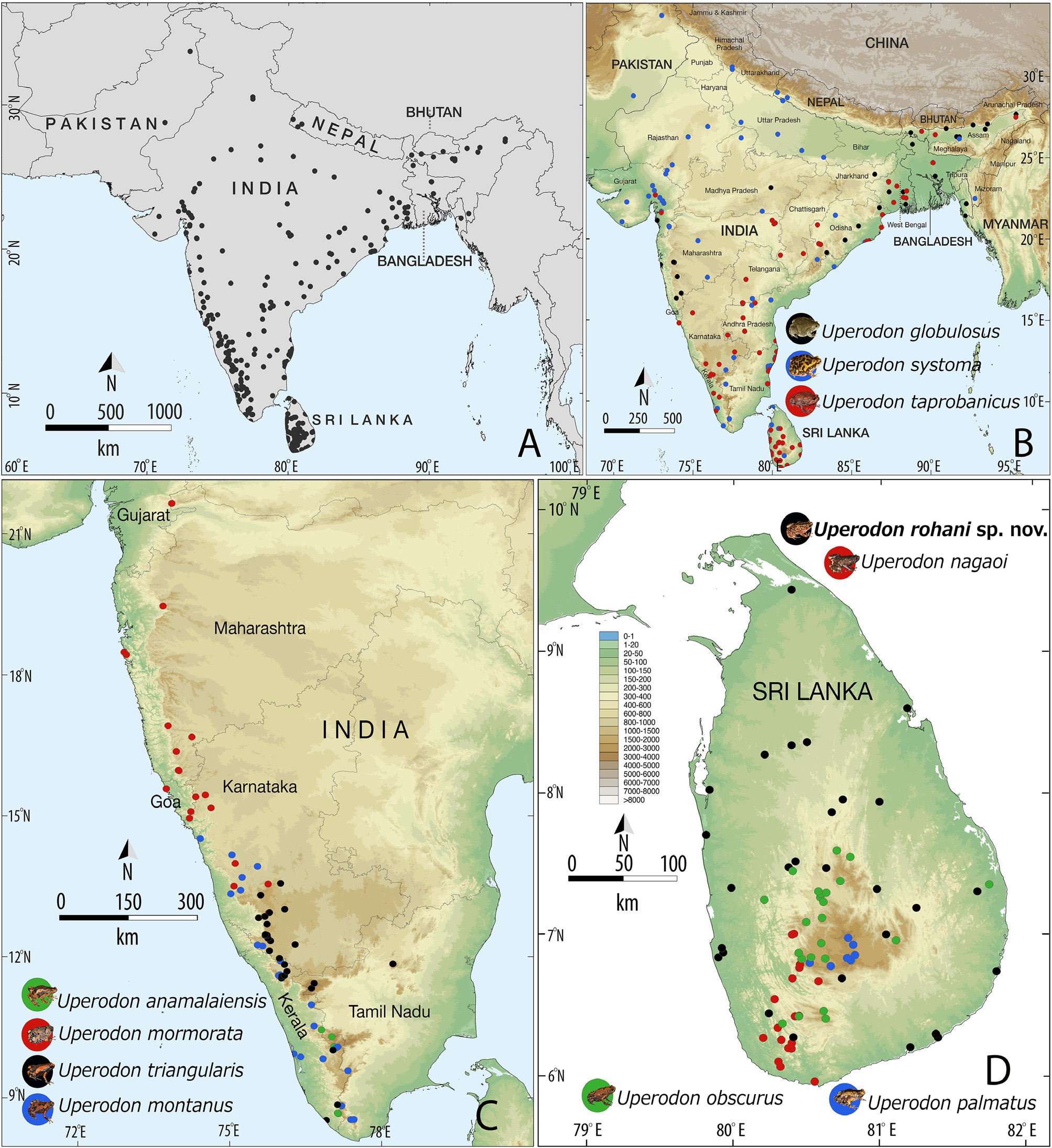
Geographical distribution and habitat. Uperodon rohani sp. nov. is endemic to Sri Lanka, where it is widely distributed throughout the island ( Fig. 2D View FIGURE 2 ). In the present study, we genetically and/or morphologically confirmed the occurrence of this species at several localities in the Central, Eastern, Northern, North Central, North Western, Southern, and Uva provinces ( Table 1). Previous records of U. variegatus from Sri Lanka refer to this species (see supplementary Table S1 and references thereto).
Comment. This species was historically considered to be conspecific with the Indian species Uperodon variegatus and widely reported as Ramanella variegata (see Table 1 and supplementary Table S1).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.