
Moinidae Goulden, 1968
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4554.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:D1B0AB6C-0415-4EBD-AF9B-71DE56ACC8A0 |
|
DOI |
https://doi.org/10.5281/zenodo.5942830 |
|
persistent identifier |
https://treatment.plazi.org/id/03D45A3B-8A37-1D57-FF2A-F9071BBFFEF0 |
|
treatment provided by |
Plazi |
|
scientific name |
Moinidae Goulden, 1968 |
| status |
|
Family Moinidae Goulden, 1968 View in CoL View at ENA
Genus Moina Baird, 1850
Moina siamensis sp. nov.
( Figs. 1–10 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Etymology. This new species is named after Siam, the former name of Thailand, the country from which it was collected.
Type locality. A roadside temporary pond in Nong Yasai district, Suphan Buri province, Thailand, (14°45'55.30"N, 99°46'05.34"E). From this locality, 60 parthenogenetic females, 60 males and 40 ephippial females were collected by Nukul Saengphan on 26 April, 2007. At the time of sampling, physicochemical parameters of water were: temperature 30.7°C, pH 7.3, conductivity 98 µs cm -1, and dissolved oxygen 5.07 mg L -1.
Holotype. A parthenogenetic female in 95% ethanol deposited at the collection of Zoological Museum of Moscow State University , MGU Ml 164 . Holotype label: “ Moina siamensis sp. nov., 1 parth. ♀ from a roadside pond, Nong Yasai district , Suphan Buri province, Thailand, HOLOTYPE ” .
Allotype. Adult male in 95% ethanol deposited at the collection of Zoological Museum of Moscow State University, MGU Ml 165 . Allotype label: “ Moina siamensis sp. nov., 1 ³ from a roadside pond, Nong Yasai district , Suphan Buri province, Thailand, ALLOTYPE ” .
Paratypes from a roadside pond, Nong Yasai district , Suphan Buri province, Thailand, preserved in 95% ethanol: 10 undissected parthenogenetic females (MGU Ml 166); 5 undissected males (MGU Ml 167); 5 undissected ephippial females (MGU Ml 168); 10 undissected parthenogenetic females (AAK 2018-060); 5 undissected males (AAK 2018-061) .
Other material studied. Twenty parthenogenetic females from temporary pools in Muang district, Suphan Buri province, 14°29'43.44"N, 99°58'40.44"E. 20 parthenogenetic females, 20 males and 20 ephippial females from Don Chedi district, Suphan Buri province, 14°38'39.48"N, 99°52'11.64"E. 30 parthenogenetic females, 10 males and 10 ephippial females, Sri Prachan district, Suphan Buri province, 14°36'33.12"N, 100°57'21.6"E, coll. 26 April 2007, Nukul Saengphan (see other localities in Table 1).
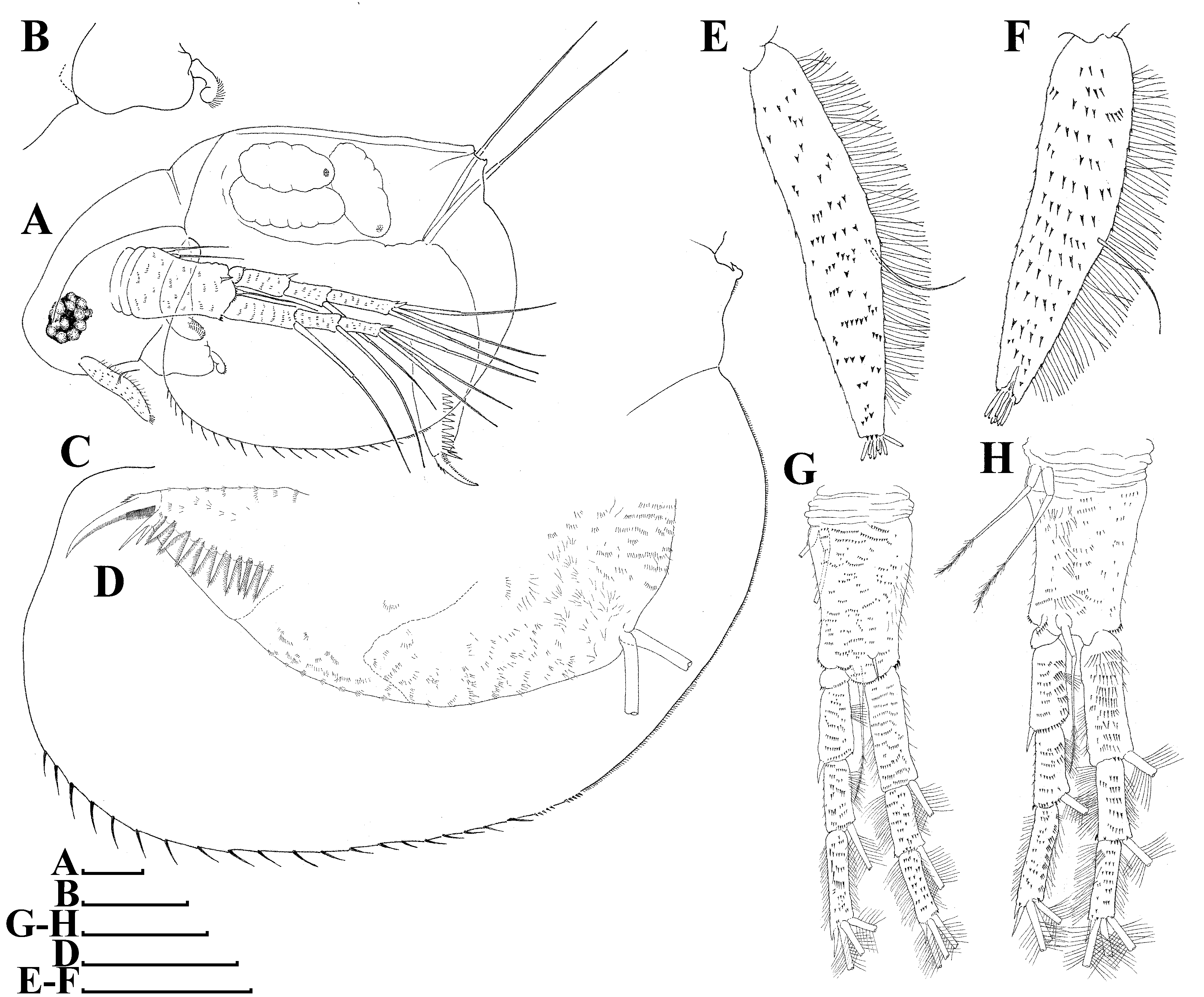
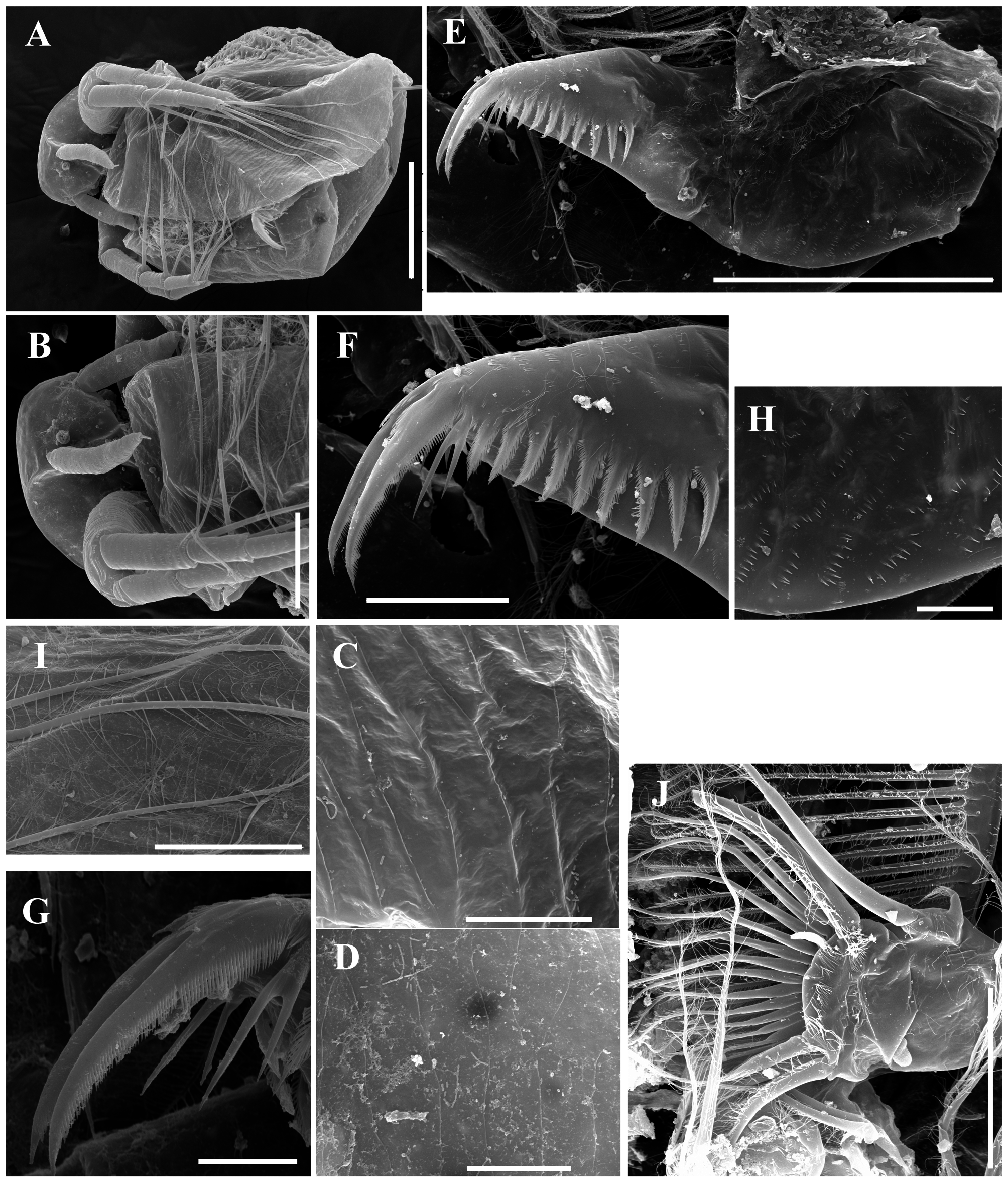
Description. Parthenogenetic female ( Figs. 1 View FIGURE 1 , 5 View FIGURE 5 A–B, E–H). General. Body ovoid in lateral view, moderately low for genus (body height/length = about 0.64), maximum height in middle portion ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 ). Dorsum of valve significantly elevated behind head, almost straight in posterior portion. Shallow dorsolateral depression separates head from rest of body. Posterodorsal angle prominent, acute, posteroventral angle broadly rounded. Ventral margin moderately convex. Anterodorsal angle prominent, rounded ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 ). Very fine sculpture on valves, forming cells elongated in dorsoventral direction ( Figs. 5 View FIGURE 5 A–B). No integumental hairs on head and valves ( Figs. 1A View FIGURE 1 , 5 View FIGURE 5 A–B, 5G–H). Body laterally compressed in anterior view. Body elongated, subovoid in dorsal and ventral view.
Head relatively large, with shallow supra-ocular depression and massive compound eye ( Figs. 1A View FIGURE 1 , 5 View FIGURE 5 A–B, G). Ocellus absent. Head pores absent.
Labrum with fleshy main body, its ventral margin slightly concave, labral plate densely setulated ( Fig. 1B View FIGURE 1 ).
Valves large, broadly ovoid ( Figs. 1A, C View FIGURE 1 , 5 View FIGURE 5 A–B). Anterior portion of ventral margin provided with moderately long setae covered by tiny setulae ( Fig. 1C View FIGURE 1 ); in posterior portion of ventral margin, long setae replaced by fine setulae. Anteriormost setulae organized in 2 or 3 groups, but then setulae gradually decreasing in size towards dorsal margin. Setulated hooks located in the dorsal most portion of posterior margin in both valves ( Fig. 1C View FIGURE 1 ).
Thorax relatively long ( Fig. 1A View FIGURE 1 ).
Abdomen short ( Fig. 1A View FIGURE 1 ).
Postabdomen elongated, conically narrowing distally ( Figs. 1A, D View FIGURE 1 , 5 View FIGURE 5 E–F); ventral margin almost straight, with rows of minute setulae ( Fig. 1D View FIGURE 1 ). Preanal margin long, convex, gradually passing to anal margin. Preanal and anal portions are covered by rows of minute setulae ( Fig. 1D View FIGURE 1 ). Postanal portion conical, both distal margin and dorsodistal angles not expressed. Laterally postanal portion of postabdomen bearing a row of 8–10 large, triangular, plumose teeth ( Figs. 1D View FIGURE 1 , 5 View FIGURE 5 E–F). Anteriormost tooth bidentate, with branches unequal in length ( Figs. 1D View FIGURE 1 , 5 View FIGURE 5 E–F).
Postabdominal setae slightly longer than postabdomen, its distal segment 2× longer than proximal segment ( Figs. 1A, D View FIGURE 1 ).
Postabdominal claw large, slightly curved, with pointed tip ( Figs. 1A, D View FIGURE 1 , 5 View FIGURE 5 E–F); its outer lateral side covered by row of denticles decreasing in size towards tip. Prominent group of larger denticles near base of claw. Ventral margin of claw with several denticles in its basal portion.
Antenna I thin (length approximately equal to 6 diameters of antennular body base), long, slightly curved ( Figs. 1 View FIGURE 1 E–F, 5G–H); surface covered by numerous fine long setae and rows of minute denticles. Antennular sensory seta slender, arising almost at middle of antennular body; 9 short aesthetascs almost subequal in size ( Figs. 1 View FIGURE 1 E–F, 5G–H).
Antenna II large and long; coxal part with 3 setulated sensory setae subequal in length ( Figs. 1 View FIGURE 1 G–H). Basal segment robust, with short distal spine on outer surface between antennal branches and long seta on inner surface; this segment covered by numerous transverse rows of fine denticles and long setae ( Figs. 1 View FIGURE 1 G–H). Antennal branches elongated. Exopod 4-segmented, subequal in size to 3-segmented endopod. All branch segments cylindrical, with rows of small denticles and setae. Antennal formula: setae 0-0-1-3/1-1-3, spines 0-1-0-1/0-0-1. Lateral and apical swimming setae of both antennal branches covered by long, fine setulae. Spine on second exopod segment short, comparable in length with both apical exopod and endopod spines ( Figs. 1 View FIGURE 1 G–H).
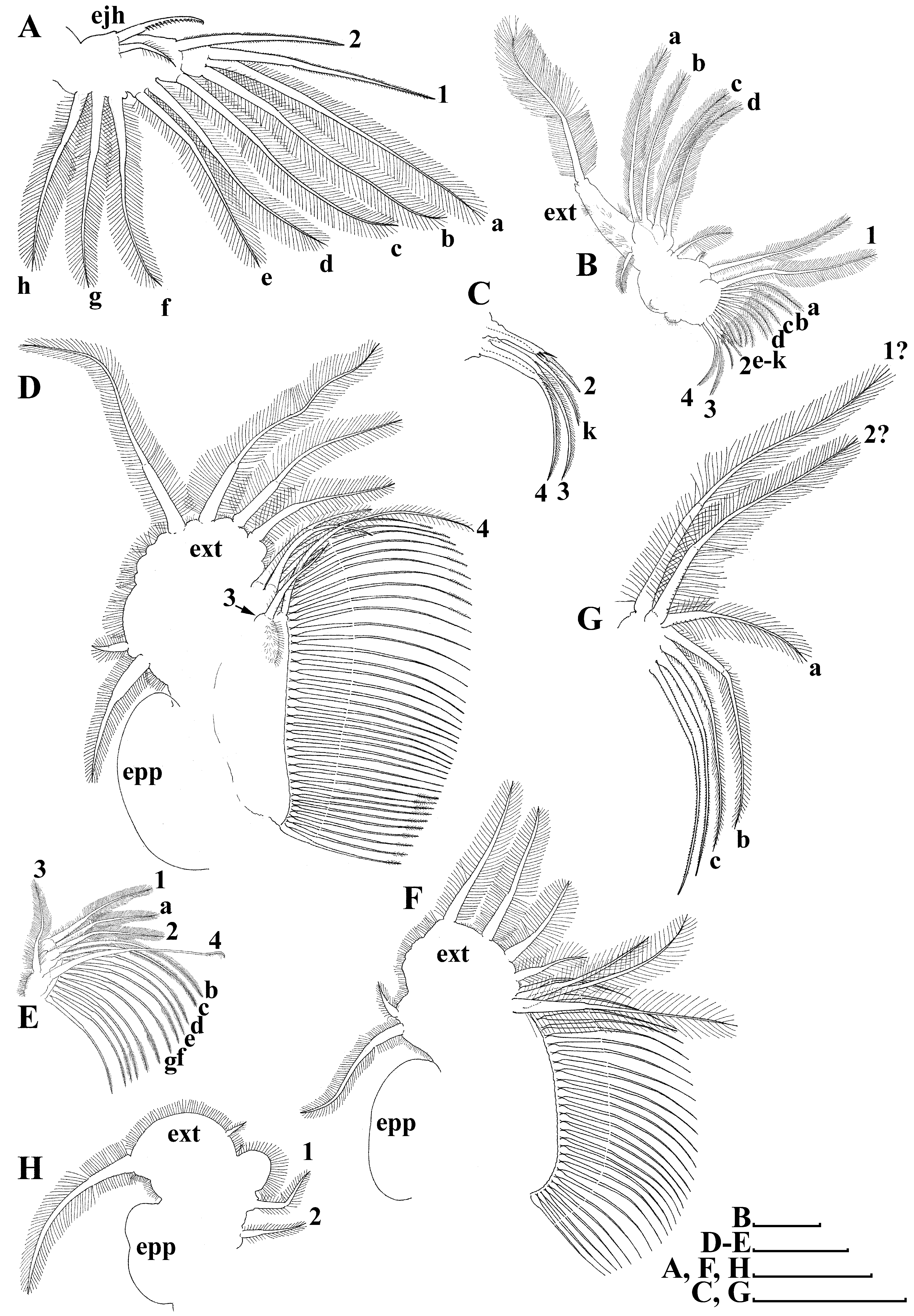
Thoracic limbs: 5 pairs ( Figs. 2 View FIGURE 2 A–H).
Limb I ( Fig. 2A View FIGURE 2 ) with elongated, narrow corm; inner distal lobe (or endite 5 sensu Kotov (2013)), with single anterior seta, bearing short setulae, and 2 soft setae ( Fig. 2A View FIGURE 2 : 1, a–b). Armature of posterior soft setae similar in all limbs: both segments provided with fine, long setulae. Endite 4 with single, relatively short anterior seta, and single posterior seta ( Fig. 2A View FIGURE 2 : 2, c). Endite 3 with 2 posterior setae, without anterior setae ( Fig. 2A View FIGURE 2 : d–e). Endite 2 with 3 posterior setae ( Fig. 2A View FIGURE 2 : f–h). Two ejector hooks of remarkably different size. No maxillar process (or endite 1 sensu Kotov (2013)) on limb base.
Limb II ( Figs. 2 View FIGURE 2 B–C) large. Limb distal portion (exopodite) as large lobe bearing large soft seta and small lateral seta; 3 endites bearing 4 soft posterior setae, and 2 small setae of unclear homology near gnathobase ( Fig. 2B View FIGURE 2 : a–d and other unnumbered setae). Gnathobase with 2 clear rows of setae ( Figs 2 View FIGURE 2 B–C): 4 anterior setae, single long seta (1) near beating seta, and 3 setae in basal corner of gnathobase (2–4), increasing in size distally, and 11 posterior setae (a–k) of gnathobase “filter plate”.
Limb III ( Figs. 2 View FIGURE 2 D–E) with large ovoid epipodite, and ovoid flat exopodite bearing 4 distal and 2 lateral setae remarkably different in length. Inner distal portion of limb with 3 endites ( Fig. 2E View FIGURE 2 ). Endite 3 with single posterior seta and single anterior seta ( Fig. 2E View FIGURE 2 : 1, a); endite 2 with single anterior and 2 posterior setae ( Fig. 2E View FIGURE 2 : 2, b–c); endite 1 with single anterior and 4 posterior setae ( Fig. 2E View FIGURE 2 : 3, d–g). Remainder of limb represented by single lobe, bearing numerous soft setae, covered by short setulae.
Limb IV ( Figs. 2 View FIGURE 2 F–G) with large ovoid epipodite, and ovoid flat exopodite, bearing 4 distal and 2 lateral setae remarkably different in length ( Fig. 2F View FIGURE 2 ). Inner distal portion of limb with 2 endites: endite 2 with anterior seta and posterior seta ( Fig. 2G View FIGURE 2 : 1?, a); endite 1 with anterior seta and 2 posterior setae ( Fig. 2G View FIGURE 2 : 2?, b–c). Other parts of limb are represented by single lobe, bearing numerous soft setae, covered by short setulae ( Figs. 2 View FIGURE 2 F–G).
Limb V ( Fig. 2H View FIGURE 2 ) with large ovoid epipodite and exopodite bearing large distal and small apical setae. Inner limb portion as flat, ovoid lobe, with setulated inner margin, and 2 setae unequal in size ( Fig. 2H View FIGURE 2 : 1–2).
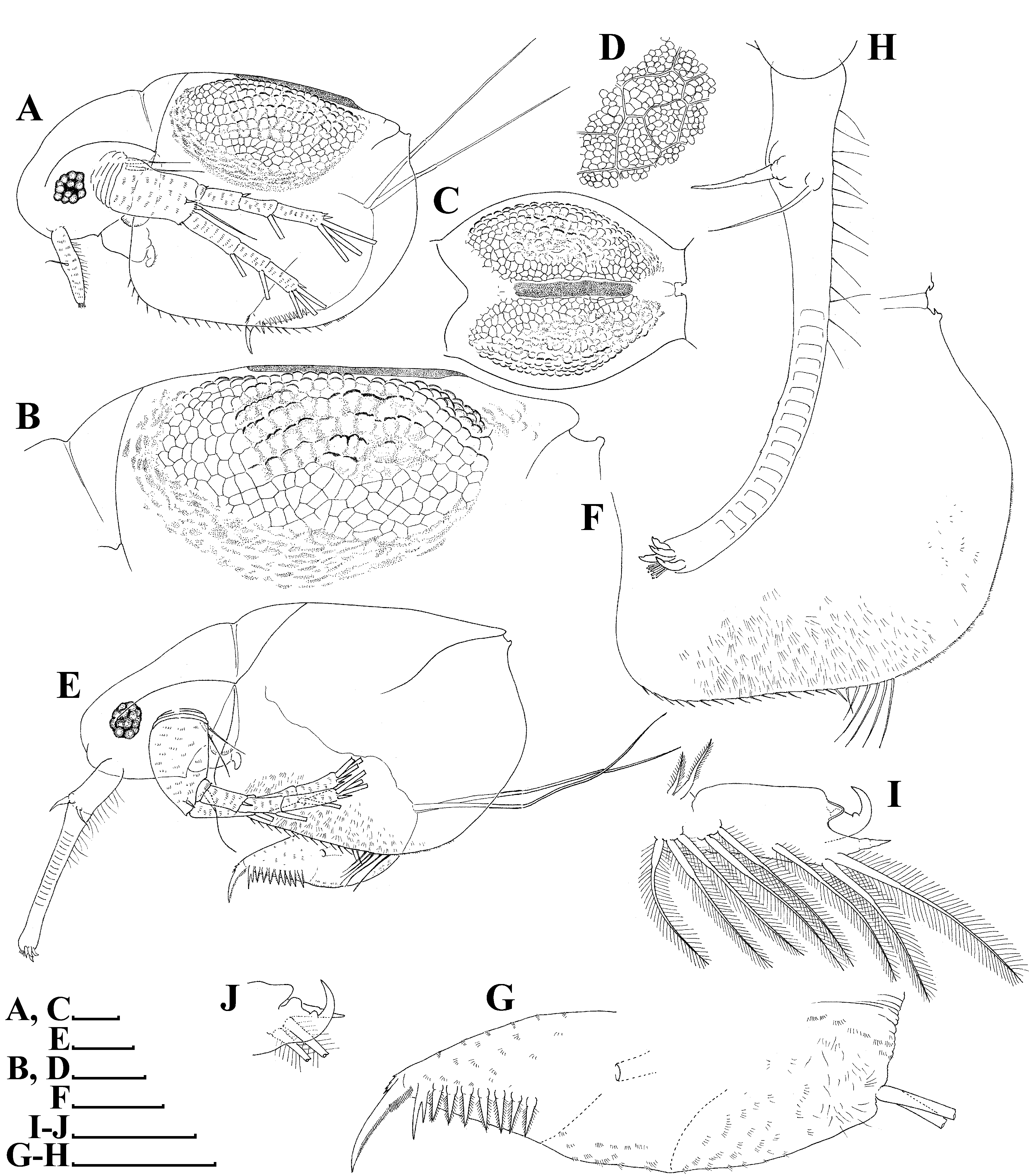
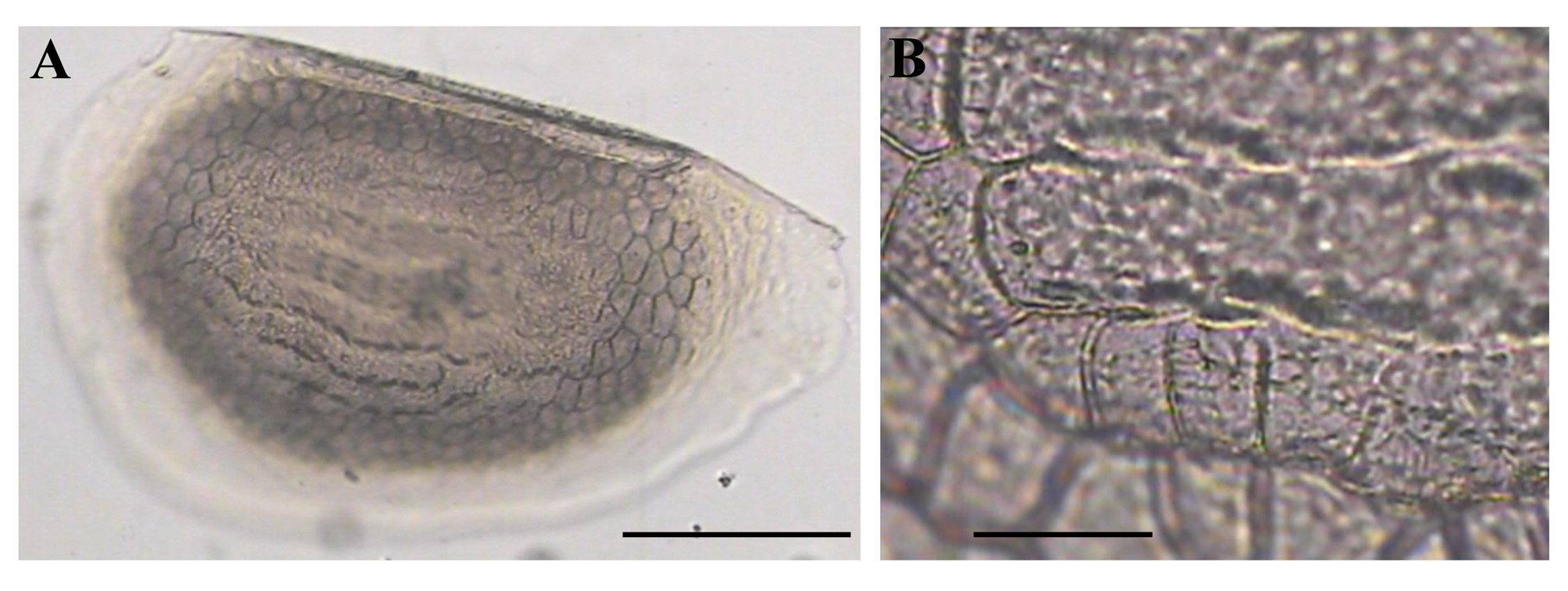
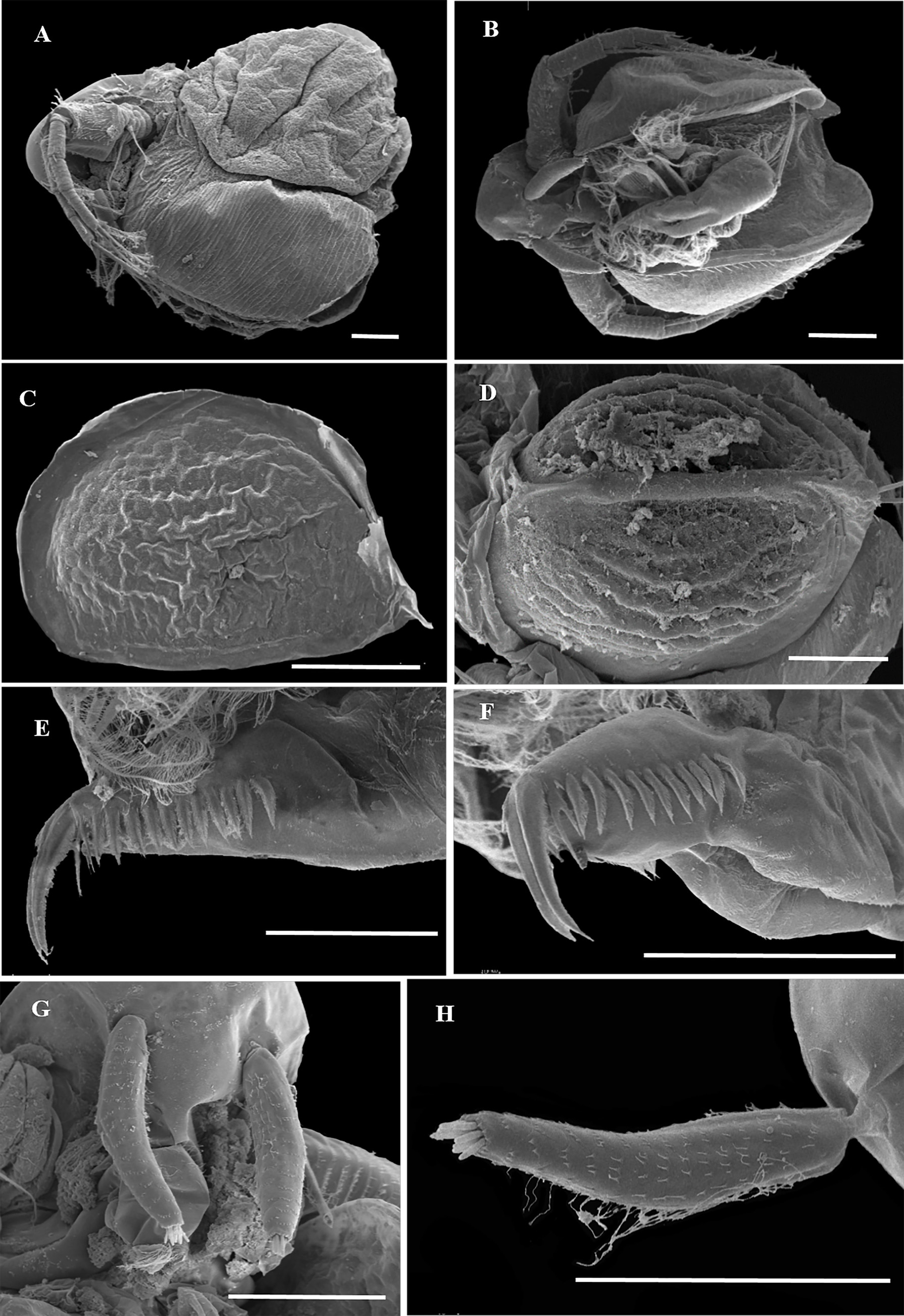
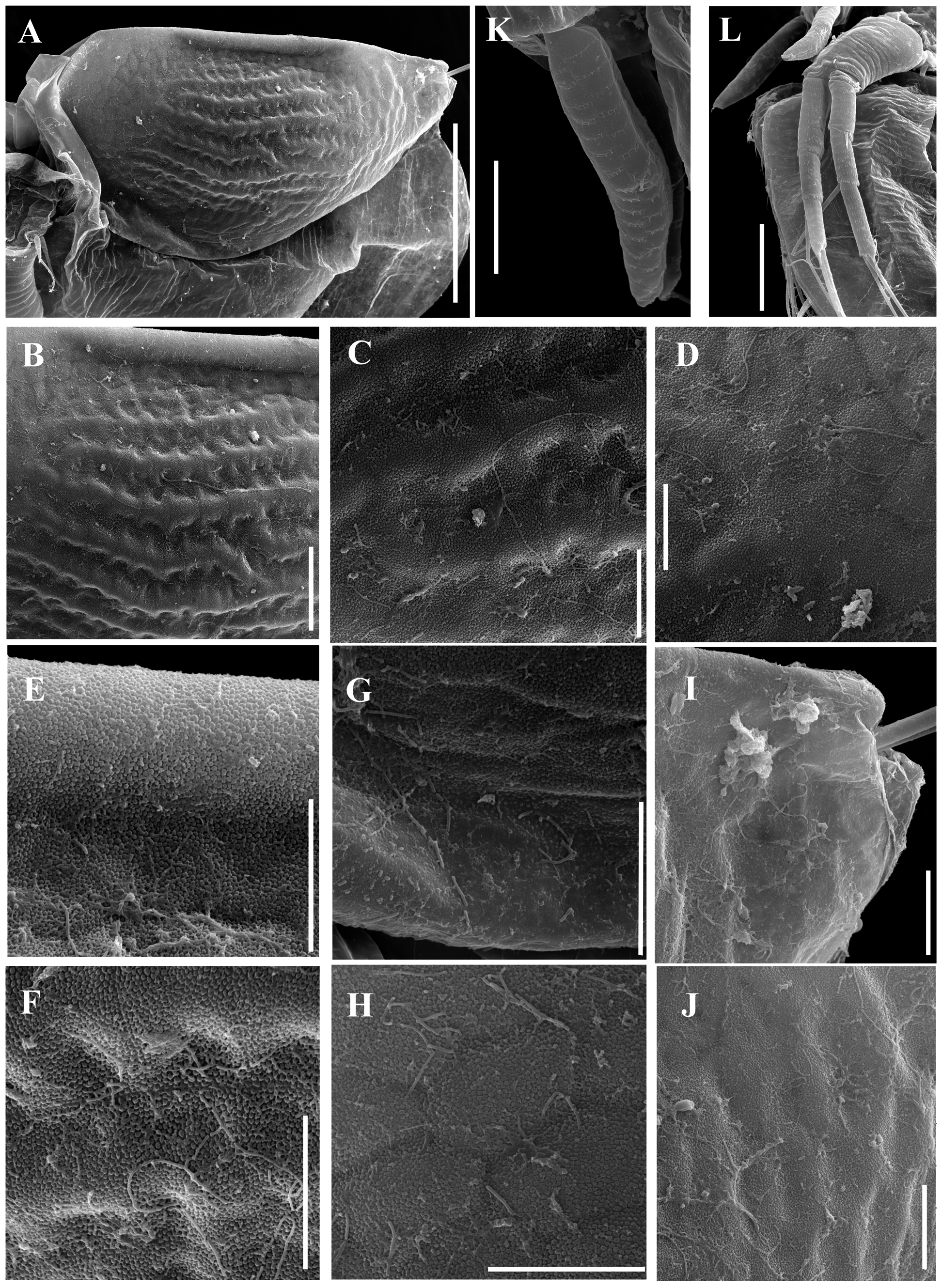
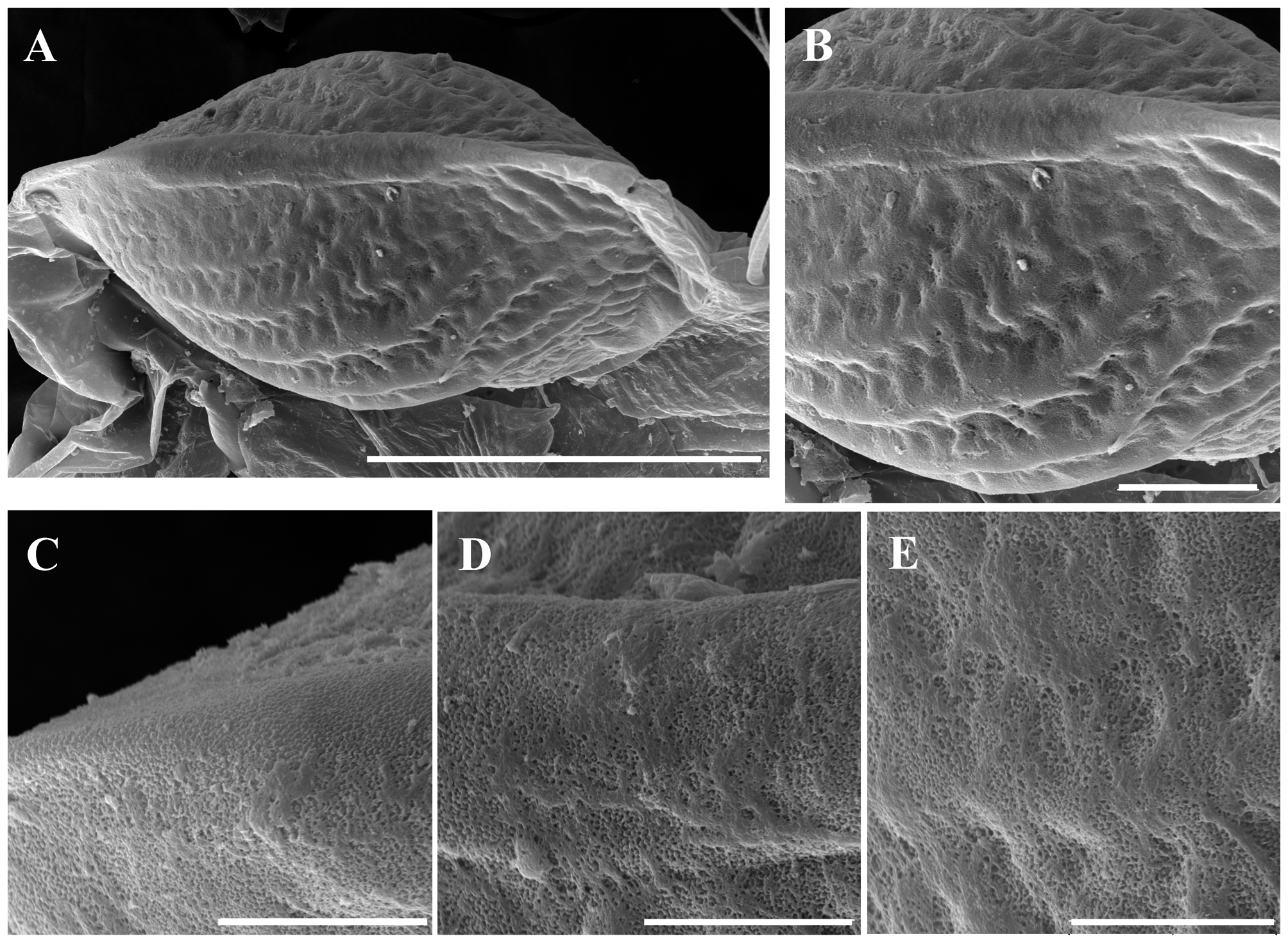
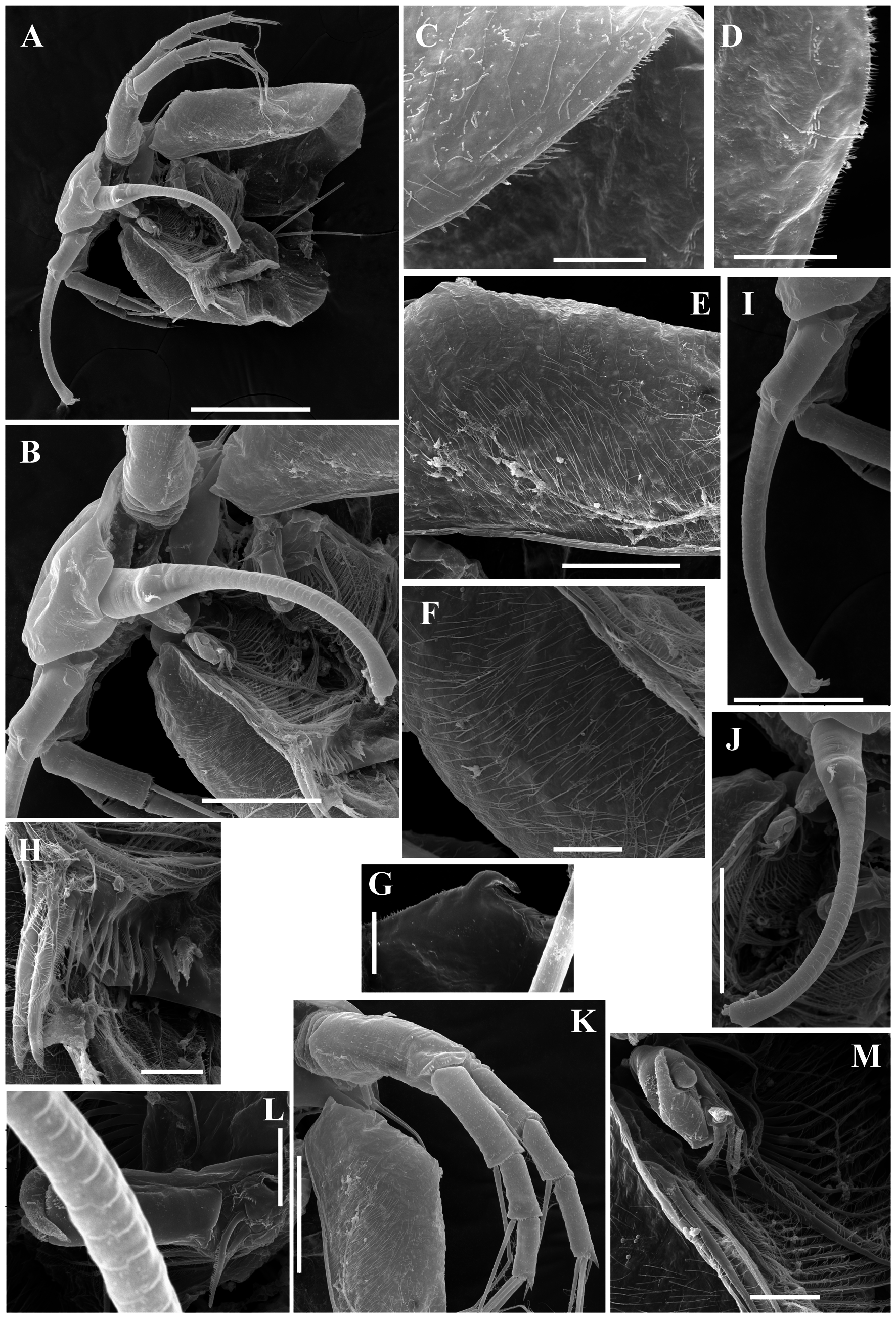
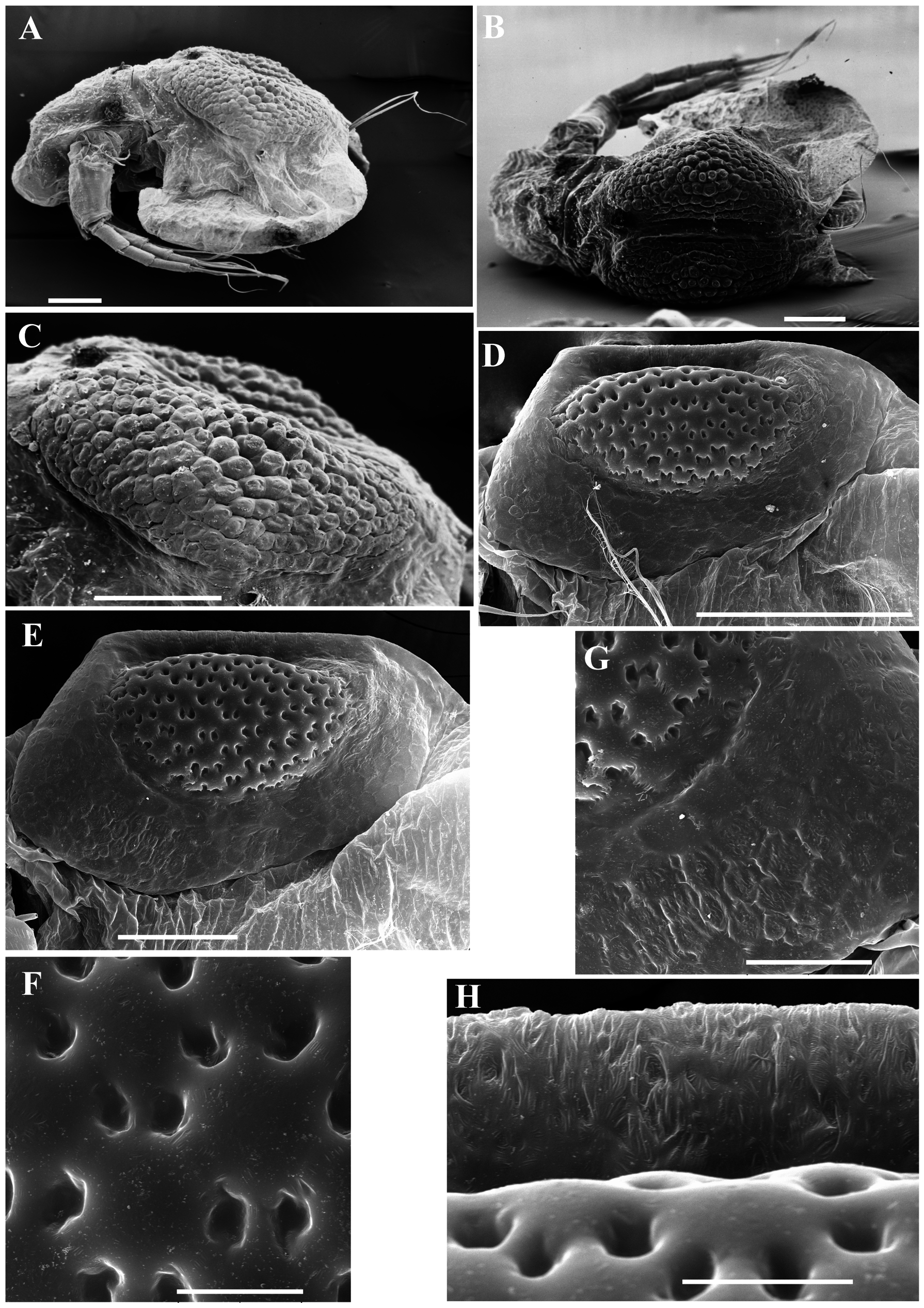
Ephippial female ( Figs. 3 View FIGURE 3 A–D, 4A, 5C–D, 6A–L, 7A–J, 8A–E). Body shape and appendages of ephippial female appearing similar to those of parthenogenetic female but with dorsal portion transformed into ephippium (for resting egg), laterally expanded (e.g. Figs. 3 View FIGURE 3 A–D). Dorsal part of valves with reinforced dorsal chitinous plate ( Figs. 3 View FIGURE 3 A–C, 4A, 5C–D, 6A–B, 8A–B), expanded to center in dorsal view. Ephippium yellow-brownish ( Figs. 4 View FIGURE 4 A– B). Macrosculpture of ephippium as polygonal cells at margins and prominent longitudinal lines in central portion ( Figs. 3 View FIGURE 3 A–D, 4A–B, 5C–D, 6H and others), these features well recognizable both under light and scanning electron microscopy. Microsculpture as tiny rounded hillocks, visible only under scanning electron microscope ( Figs. 6 View FIGURE 6 B–J, 8B–E). Ephippium containing single resting egg.
Male ( Figs. 3 View FIGURE 3 E–J, 9A–E, 10A–M). General. In lateral view body ovoid, more elongated compared to female (body height/length about 0.55) ( Figs. 3E View FIGURE 3 , 9 View FIGURE 9 A–B). Dorsal margin of valves slightly elevated above head, posteroventral angle distinct ( Figs. 3E View FIGURE 3 , 9 View FIGURE 9 A–B).
Head longer than in female; labrum somewhat thinner than that of females, with large, setulated distal labral plate ( Figs. 3E View FIGURE 3 , 9 View FIGURE 9 A–B, 10B). Head pores absent. Compound eye large. Ocellus absent.
Valve ovoid, more elongated than that in female, with short setae, decreasing in size posteriorly, along ventral margin ( Figs. 3E View FIGURE 3 , 9 View FIGURE 9 A–B, 3F). Setulated spine on inner face of valve near posteroventral angle. Posterior portion of ventral margin with successive series of fine, exactly marginal setulae. Anterior portion of ventral margin with row of setae, increasing in size to central part with 4 or 5 setae more than 6× longer than those of anterior part. In contrast to parthenogenetic and gamogenetic females, outer surface of valves in males covered by fine relatively long hairs ( Figs. 3F View FIGURE 3 , 9C View FIGURE 9 , 10 View FIGURE 10 C–G).
Thorax ( Fig. 3E View FIGURE 3 ) relatively long.
Abdomen ( Fig. 3E View FIGURE 3 ) short.
Postabdomen generally as in female, with large bidentate tooth distally (distal branch always significantly large than basal branch), and row of 7–9 large, triangular plumose teeth ( Figs. 3E, G View FIGURE 3 , 10H View FIGURE 10 ). Gonopore opening in lateral surface of postabdomen.
Postabdominal setae ( Fig. 3E View FIGURE 3 ) almost 2× longer than postabdomen. Its distal segment significantly longer than proximal one.
Antenna I very long and regularly curved, covered by tiny hairs and rows of small denticles ( Figs. 3H View FIGURE 3 , 9 View FIGURE 9 C–E, 10I–J). Antennular sensory seta long, arising from first quarter of antennular body. Male seta more robust, located near sensory seta. Apical tip of antennular body separated into 2 parts: first with 9 short aesthetascs, second with 4 hooks.
Thoracic limb I essentially similar to that in parthenogenetic female, but with large, curved copulatory hook with setulated margin and pointed apex ( Figs. 3 View FIGURE 3 I–J, 10L–M).
Size. Adult parthenogenetic females 0.50–0.99 mm in length; ephippial females up to 0.92 mm in length; adult males 0.50–0.89 mm in length. Juvenile females to 0.5 mm in length. Holotype 0.99 mm in length; allotype 0.89 mm in length.
Variability. No significant variability was found in individuals between localities.
Differential diagnosis. In tropical Asia, Moina siamensis sp. nov. is close to M. weismanni Ishikawa, 1896 , resulting in possible confusion. Morphology of parthenogenetic females and males of both species is nearly identical. Based on current knowledge of moinid morphology, we suggest that the ephippium should be used as the primary and most reliable diagnostic feature for discrimination between sibling species. The mature ephippium of M. siamensis sp. nov. is characterized by prominent longitudinal lines in its central region, whereas the ephippium of M. weismanni ( Figs. 12 View FIGURE 12 A–C) is strongly tuberous, with distinct borders between tubercles. These features are recognizable both under light and scanning electron microscopes. However, we can only identify populations that contain mature gamogenetic females. Primary differences between Moina siamensis sp. nov. and other tropical moinids are discussed below.
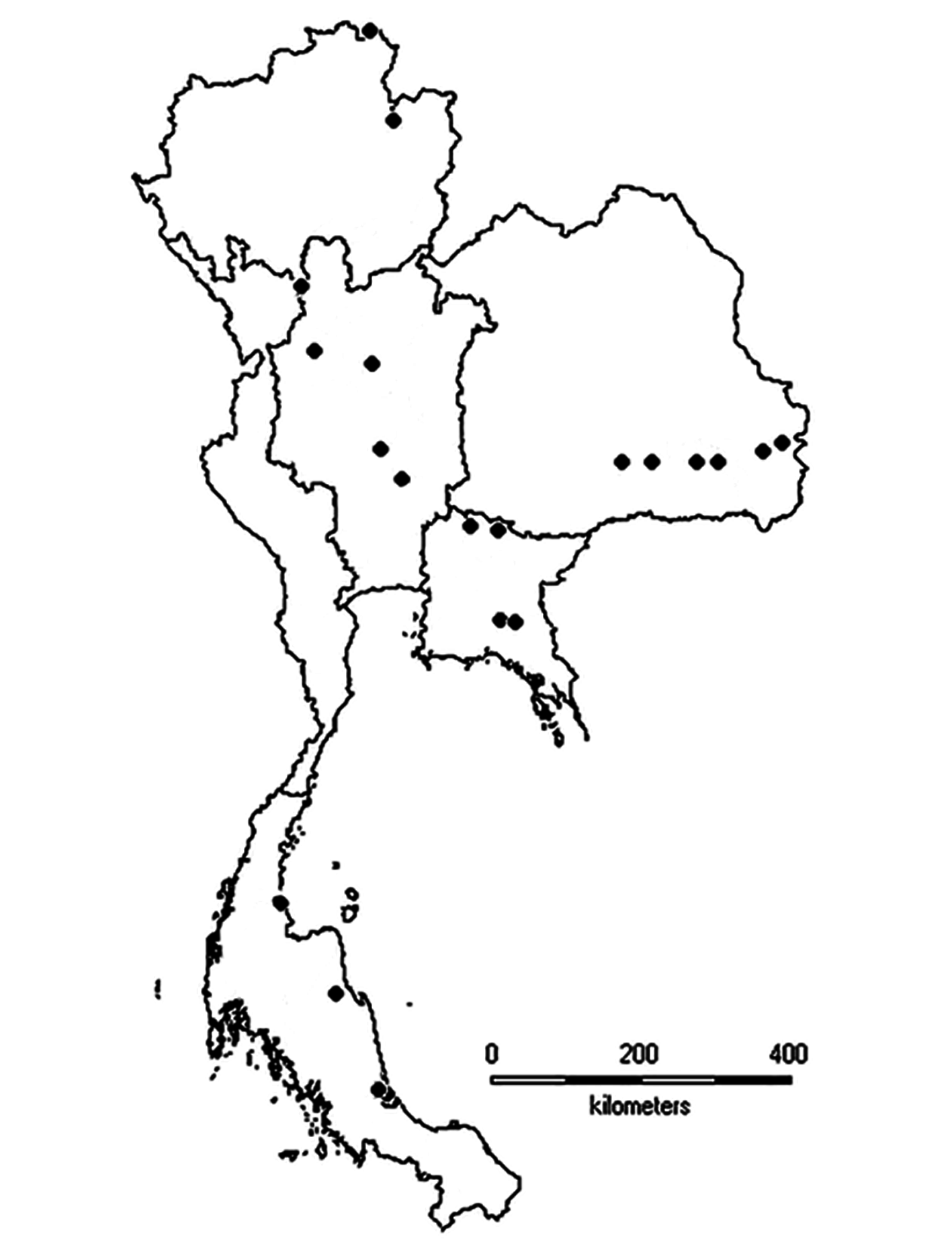
Distribution and ecology. Until now, Moina siamensis sp. nov. was known only from temporary water bodies in Thailand ( Table 1, Fig. 11 View FIGURE 11 ). It occurs nationwide, including northern, central, northeastern, eastern, and southern regions. This species inhabits shallow, roadside ponds, and pools (especially rice fields) with developed vegetation buffers, industrial wastewater treatment ponds, and animal farms. Moina siamensis sp. nov. was collected during the wet season (May and October). Some physicochemical parameters of the water were: temperature 26.5–34.0°C, conductivity 48.0–142.0 µScm -1, pH 6.1–8.5, and dissolved oxygen 1.75–7.82 mg L -1. Moina siamensis sp. nov. usually co-exists with M. micrura , but the latter species occurs in both temporary and permanent ponds ( Sanoamuang 1998).
Economical value for aquaculture. The sizes of adults and juveniles of M. siamensis sp. nov. range from 0.25–0.50 and 0.50–0.99 mm, which are suitable to feed juvenile freshwater fish or shrimps. Recent culture experiments demonstrated that M. siamensis sp. nov. contains 61.04% protein and 7.96% lipids. They can be cultured commercially in Thailand. Adults and dried eggs of this new species are presently sold in local markets.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |