
Thyroptera wynneae, Velazco & Gregorin & Voss & Simmons, 2014
|
publication ID |
https://doi.org/ 10.1206/3795.1 |
|
publication LSID |
lsid:zoobank.org:pub:9EA6163C-7A05-4550-B222-8C5F37BDFB5D |
|
persistent identifier |
https://treatment.plazi.org/id/2C830ED6-FC2C-40D8-96D8-92A0187E2B77 |
|
taxon LSID |
lsid:zoobank.org:act:2C830ED6-FC2C-40D8-96D8-92A0187E2B77 |
|
treatment provided by |
Carolina |
|
scientific name |
Thyroptera wynneae |
| status |
sp. nov. |
Thyroptera wynneae View in CoL , new species
Patricia’s Disk-winged Bat
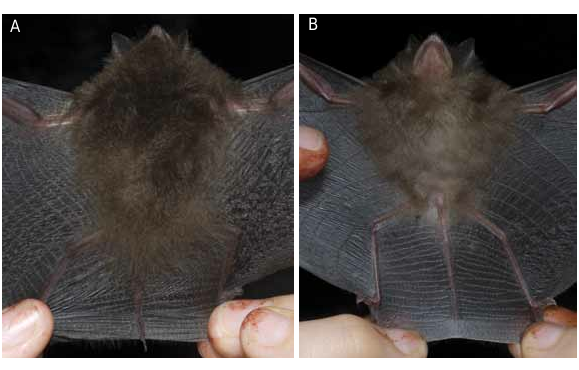
HOLOTYPE: The holotype (CEBIOMAS 237; figs. 1 View FIG , 2 View FIG , 4E View FIG , 5E View FIG , 6E View FIG ), an adult male specimen preserved in alcohol with the skull removed and cleaned, was collected by Robert S. Voss and subsequently prepared by Paúl M. Velazco (original number: PMV 2469 View Materials ) on 29 January 2012 at the Centro de Investigaciones Jenaro Herrera (45°05′S, 73°40′W), a forestry research station located about 2.5 km inland from the right bank of Río Ucayali in the Peruvian department of Loreto ( fig. 3 View FIG , locality 27). Frozen tissues are deposited at the American Museum of Natural History ( AMNH 278486 About AMNH ). GoogleMaps
PARATYPES: Two additional specimens ( CMUFLA 694 and CMUFLA 1160 ), both adult males preserved in alcohol with the skulls removed and cleaned, were collected at a site called Campolina (ca. 19°42′S, 42°30′W; fig. 3 View FIG ), about 25 km E of Marliéria in the Parque Estadual do Rio Doce , Minas Gerais, Brazil. Campolina is shown on a map of bat sampling localities by Tavares et al. (2007: fig. 1 View FIG ), who also worked at this locality. One paratype ( CMUFLA 1160 ) was collected by Renato Gregorin on 11 January 2013 and the other ( CMUFLA 694 ) was collected by Karina Lobão Vasconcellos and Bruno Bret Gil (original field number: PERD 310 ) on 11 March 2013. CMUFLA 694 has a damaged skull (the braincase is broken in the parietal region and the left auditory bulla is missing), but the other specimen is in good condition GoogleMaps .
DISTRIBUTION: Thyroptera wynneae is currently known only from the type locality in northeastern Peru and from the Parque Estadual do Rio Doce in southeastern Brazil ( fig. 3 View FIG ; appendix 1) .
DIAGNOSIS AND DESCRIPTION: Thyroptera wynneae is a small thyropterid with a short, slender rostrum and a globular braincase ( figs. 1–2 View FIG View FIG , 4–6 View FIG View FIG View FIG ). The dorsal fur is light brown with a slightly grayish overtone. Dorsal hairs are unicolored for most of their length, but a tiny basal portion of each hair is lighter than the rest of the shaft ( fig. 1A View FIG , 2A View FIG ). Ventral hairs are tricolored with the basal third of each hair whitish in color, the middle segment light brown, and the tip darker. The fur is long and woolly, approximately 11 mm long between the shoulders and 7 mm on the chest. The proximal portion of the forearm (roughly 12 mm of the shaft just distal to the elbow) is densely furred with long hairs. Most of the dorsal surface of the pinna is densely covered with long hairs, but the distal portion is naked ( fig. 1A View FIG ).
The wing membranes of Thyroptera wynneae are dark brown. The plagiopatagium and uropatagium ( fig. 1C View FIG , 2A View FIG ) are marked by transverse lines of elastine fibers punctuated by regularly spaced wartlike somatosensory domes (sensu Zook, 2007). The dorsal surface of the uropatagium is densely covered with long hairs proximally, and sparsely covered with long hairs distally. The tip of the wing is also sparsely covered with long hairs, and the uropatagium, plagiopatagium, and dactilopatagium have hairs distributed sparsely along their edges. The adhesive disks of the thumb and foot are oblong ( fig. 1B, 1C View FIG ). The thumb disks vary somewhat in size among individuals, ranging from 2.7–4.0 mm in length and 1.8–3.1 mm in width (length being measured along the longest axis of the disk). Similarly, foot disks range from 1.6–2.5 mm in length and 1.4–2.2 mm in width. Some of this variation, however, might be due to differences in preservation. The calcar is long (8.5–12.0 mm) and has two lappets (small cartilaginous flanges) that project posterolaterally from the shaft of the calcar. In addition to these skeletal processes, five tiny skin projections are also present between the foot disk and the proximal lappet ( fig. 1C View FIG ).
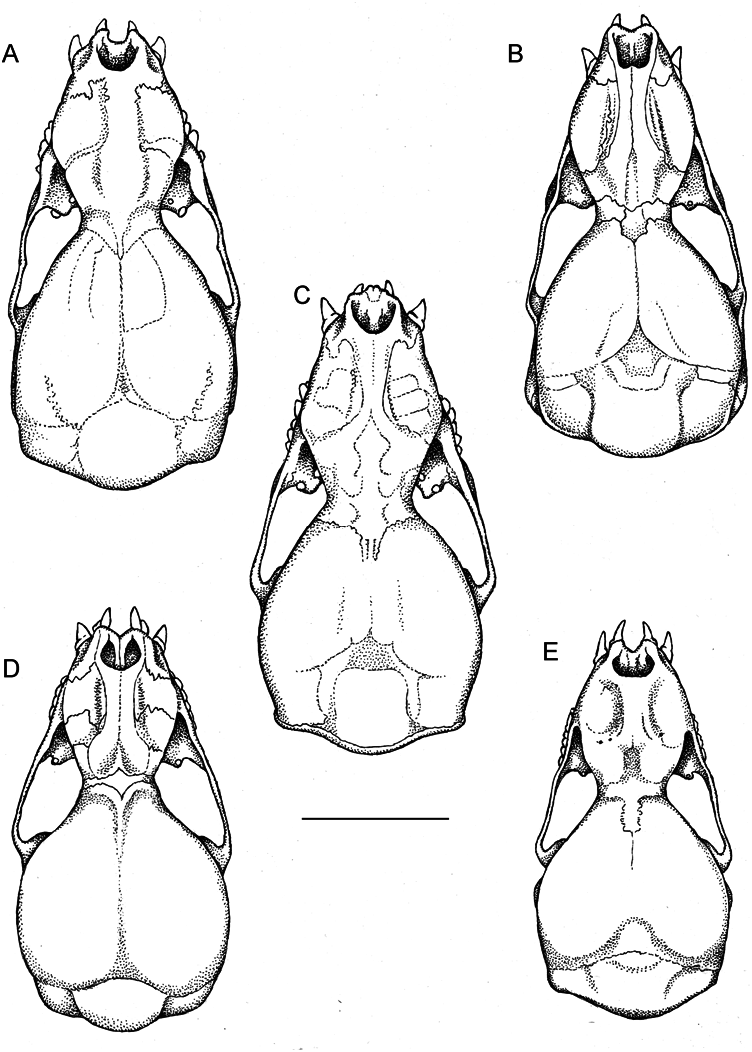
The skull of Thyroptera wynneae has a smoothly rounded braincase that lacks a sagittal crest ( figs. 4E View FIG , 6E View FIG ). The angle between the rostrum and braincase is approximately 130º ( fig. 6E View FIG ) and the rostrum is considerably shorter than the braincase ( table 1). There is a large foramen ovale which is 0.5 mm in length. A transverse ridge is present between the right and left hamular processes of the pterygoid. This ridge is intersected anteriorly by a midventral bladelike crest, dividing the mesopterygoid fossa into two separate concavities ( fig. 5E View FIG ; Gregorin et al., 2006, fig. 2A View FIG ).
Like all other thyropterids, Thyroptera wynneae has a dental formula of I2/3, C1/1, P3/3. M3/3 = 38 teeth. The upper incisors are bicuspidate. The second upper incisor (I2) has a mesial cusp that is larger than the distal cusp. Both cusps on I2 are perpendicularly arranged relative to the long axis of the toothrow. The first and second upper incisors are in contact with one other and are separated from the canine by a small diastema. P1 is rounded in occlusal view. Anteroposterior length of P1 is less than that of P4, and crown height of P1 is slightly less than that of P4. P1 and P4 both have narrow lingual and labial cingula. P4 and P5 are subequal in anteroposterior length but crown height of P4 is slightly less than that of P5. P5 has a narrow labial cingulum and a very large lingual cingulum that forms a distinct lobe, the latter extending as far lingually as the protocone of M1. A well-developed anterolingual cusp is present on the labial cingulum of P5. M1 and M2 are subequal in size. A parastyle is present and the protocone is well developed on all upper molars including M3. A metastyle is present and the paracone and metacone are subequal in size on M1 and M2. In contrast, a metastyle is absent and the paracone is taller than the metacone on M3. Lingual cingula are present on all three upper molars. The preparacrista is shorter than the postparacrista on M1 and M2, but the preparacrista is longer than the postparacrista on M3. The premetacrista is shorter than the postmetacrista on M1 and M2, but the postmetacrista is absent on M3.
The first and second lower incisors (i1 and i2) are tricuspidate. The third lower incisor (i3) has two small accessory cusps on each side (mesostyles and distostyles). The i3 is subequal in height to i1 and i2, and is 1.5 times the buccolingual width of i1 and i2. The p1 is triangular in occlusal view. The p4 and p5 are buccally flattened and ovate in occlusal view. All three lower premolars are subequal in height. The lower molars are similar in shape. The m3 is slightly shorter in anteroposterior length than m1 and m2. The protoconids are taller than the hypoconids on all three lower molars. All of the cristids on all lower molars are clearly notched.
COMPARISONS: External and craniodental measurements for Thyroptera wynneae and its congeners are provided in tables 1 and 2. T . wynneae can be easily distinguished from T. devivoi , T. lavali , and T. tricolor by its shorter forearm and smaller greatest length of skull ( tables 1–2). T. wynneae and T. discifera overlap somewhat in size ( tables 1–2) but can be unambiguously distinguished based on pelage characteristics.
Externally, Thyroptera wynneae can be distinguished from all the other species in the genus by its tricolored ventral pelage, which contrasts with the unicolored ventral pelage of T. discifera and T. tricolor , and with the bicolored ventral pelage of T. devivoi and T. lavali . The overall appearance of the ventral pelage is light brown in T. wynneae , whereas it is grayish brown in T. devivoi , yellowish brown in T. discifera , medium brown in T. lavali , and white or whitish in T. tricolor . Dorsally the hairs between the shoulders are long (11 mm) and woolly in T. wynneae ( fig. 2A View FIG ) while they are long (8.0–12.0 mm) and not woolly in T. discifera and T. tricolor , and short (3.5–6.0 mm) and woolly in T. devivoi and T. lavali . The proximal portion of the forearm is well furred in T. wynneae and T. discifera , whereas it is only sparsely haired in T. devivoi , T. lavali , and T. tricolor .
The dorsal surface of the uropatagium is densely covered with long hairs proximally, and sparsely covered distally, in T. wynneae and T. discifera . In contrast, the entire dorsal surface of the uropatagium is sparsely covered with short hairs in T. lavali and T. tricolor . The tip of the wing is covered sparsely by long hairs (> 1.8 mm) in T. wynneae and T. tricolor , whereas these hairs are short (<1.2 mm) in T. discifera and T. lavali (we were not able to examine T. devivoi for this characteristic). The adhesive disks of the thumb are oblong in T. wynneae ( fig. 1B View FIG ), T. devivoi , and T. lavali , whereas they are circular in T. discifera and T. tricolor . The calcar has two lappets that project posterolaterally from the shaft of the calcar in T. wynneae ( fig. 1C View FIG ) and T. tricolor . In contrast, they are absent or only one faintly developed lappet is present in T. devivoi , and only a single lappet is present in T. discifera and T. lavali . Five tiny skin projections are present between the foot disk and the proximal lappet in T. wynneae ( fig. 1C View FIG ) and T. tricolor , whereas these structures are entirely absent in T. devivoi , T. discifera , and T. lavali .
Cranially, Thyroptera wynneae can be distinguished from all other species of Thyroptera by having a rostrum that is considerably shorter than the braincase. The rostrum is of equal length or longer than the braincase in T. devivoi , T. discifera , T. lavali , and T. tricolor . The foramen ovale is large (0.50–0.65 mm) in T. wynneae , T. devivoi , T. lavali , and T. tricolor , whereas it is smaller (<0.45 mm) in T. discifera . The transverse ridge present between the hamular processes of the pterygoid is anteriorly intersected by a midventral bladelike crest, dividing the mesopterygoid fossa in two separate concavities in T. wynneae , T. devivoi , T. lavali , and T. tricolor . The midventral bladelike crest does not reach the transverse ridge in T. discifera ( fig. 5 View FIG ).
Dental characters can also be used to distinguish species of Thyroptera . The mesial cusp is larger than the distal cusp on I 2 in T. wynneae , T. discifera , and T. tricolor , whereas these cusps are subequal in T. devivoi and T. lavali . Both cusps on I2 are perpendicularly arranged relative to the long axis of the toothrow in T. wynneae , T. devivoi , and T. lavali , whereas they are obliquely arranged in T. discifera and T. tricolor . The first and second upper incisors are in contact with each other in T. wynneae , T. devivoi , T. lavali , and T. tricolor but are separated by a small diastema in T. discifera . P1 is rounded in occlusal view in T. wynneae , T. devivoi , T. lavali , and T. tricolor , whereas it is elongated in T. discifera . All three lower incisors are subequal in height in T. wynneae , whereas the i3 is taller than i1 and i 2 in T. devivoi , T. discifera , T. lavali , and T. tricolor . The third lower incisor has two small accessory cusps on each side (mesostyle and distostyle) in T. wynneae and T. discifera , but these accessory cusps are only faintly developed in T. devivoi and T. tricolor and are either faintly developed or entirely absent in T. lavali . The i3 is 1.5 times the buccolingual width of i1 and i 2 in T. wynneae , T. discifera , and T. tricolor , whereas the i3 is twice the buccolingual width of i1 and i 2 in T. devivoi and T. lavali . The p4 and p5 are buccally flattened and ovate in occlusal view in T. wynneae , T. discifera , whereas they are flattened and squarish in T. tricolor , and wide and squarish in T. devivoi and T. lavali .
MULTIVARIATE ANALYSIS: Our multivariate statistical analysis included only male specimens because females are unknown for Thyroptera wynneae . We conducted a principal component analysis of measurements of 21 specimens of Thyroptera , including 3 each of T. wynneae , T. devivoi , and T. discifera , 2 of T. lavali , and 10 of T. tricolor (appendix 1). The first three principal components accounted for 88.5% of the total variance in the log-transformed measurements of this material ( table 3). PC 1 accounted for the highest percentage (70.1%), and this vector has uniformly positive loadings, suggesting that it is a size factor (with a notably high loading for forearm length). Correspondingly, specimens of small-bodied species ( T.wynneae and T. discifera ) have lower scores along this axis, large-bodied T. lavali has high scores, and medium-sized species ( T. tricolor and T. devivoi ) have intermediate values ( fig. 7 View FIG ). Although T. wynneae and T. discifera have overlapping scores on PC2 ( fig. 7A View FIG ), these species are resolved as distinct clusters on PC3 ( fig. 7B View FIG ). Factor loadings on PC3 suggest that this separation is primarily attributable to taxonomic differences in forearm length and width at M3.
NATURAL HISTORY: The Centro de Investigaciones Jenaro Herrera is a forestry research station located about 2.5 km inland from the right bank of the Río Ucayali in the Peruvian department of Loreto. The station is on a terrace above the river floodplain and is not subject to annual flooding by river water. The local vegetation consists of primary and secondary lowland rainforest (“high terrace forest” sensu López-Parodi and Freitas, 1990) on well-drained soils, but a distinctive riparian formation (“creek forest” sensu López-Parodi and Freitas, 1990; “bosque de quebrada” sensu Kahn and Mejia, 1990) occurs along streams, and palm swamps dominated by Mauritia flexuosa (locally known as “aguajales”) occupy depressions with permanently waterlogged soils. Small plantations (of yuca, pineapple, etc.) are also scattered throughout the area. The holotype of Thyroptera wynneae was found roosting with another individual (which escaped capture) in the late morning (about 11:00 hrs). Both bats were concealed within a dead Cecropia leaf that was hanging from understory vegetation by its petiole about 2 m above the ground in secondary forest next to a trail about 50 m from the main buildings of the biological station ( fig. 8 View FIG ).
The Parque Estadual do Rio Doce protects some 36,000 hectares of semideciduous forest, most of which is thought to be secondary growth ( Lopes et al., 2002). The park includes hills and low mountains ( Stallings, 1988; Lopes et al., 2002), but Campolina is located in primary forest with tall (20–30 m) trees and a relatively open understory on level terrain. Both paratypes were captured in ground-level mist nets.
ETYMOLOGY: We are pleased to name this species in honor of Patricia J. Wynne ( fig. 9 View FIG ), longtime artist-in-residence in the AMNH Department of Mammalogy, who has drawn mammals for scientific and educational publications for over 40 years. Exceptional for clarity, elegant rendering, and meticulous attention to relevant detail, Patricia’s work is immediately recognizable and inimitable (e.g., figs. 3–6 View FIG View FIG View FIG View FIG ).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |