
Campostichomma Karsch, 1892
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4362.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:61A41EC1-2557-429D-B47C-595D12F2D8E3 |
|
DOI |
https://doi.org/10.5281/zenodo.5999768 |
|
persistent identifier |
https://treatment.plazi.org/id/03D38783-FF86-FF85-E6B3-68A3FEBA617E |
|
treatment provided by |
Plazi |
|
scientific name |
Campostichomma Karsch, 1892 |
| status |
|
Campostichomma KArsch, 1892 : 296, pl. 12, FIg. 19 (TypE spEcIEs by OrIgInAl dEsIgnAtIOn: Campostichomma manicatum KArsch, 1892 ); SImOn 1898A: 7; LEhtInEn 1967: 221; GrIswOld 1993: 7; GrIswOld et al. 1999: 59; POlOtOw et al. 2015: 151 (trAnsFErrEd tO UdubIdAE View in CoL ); WSC 2017.
Note. Name is neuter in gender, hence Campostichomma manicatum .
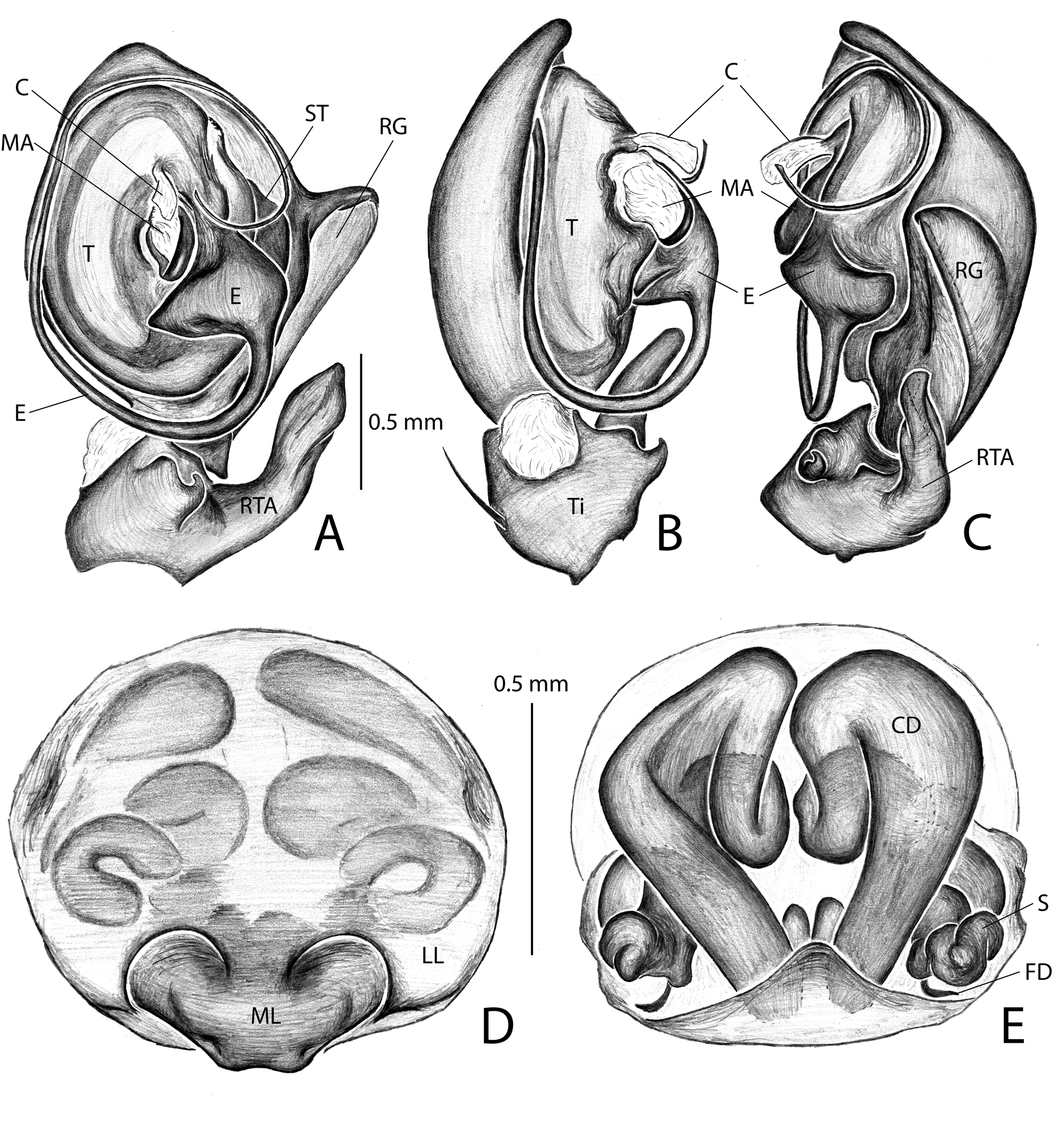
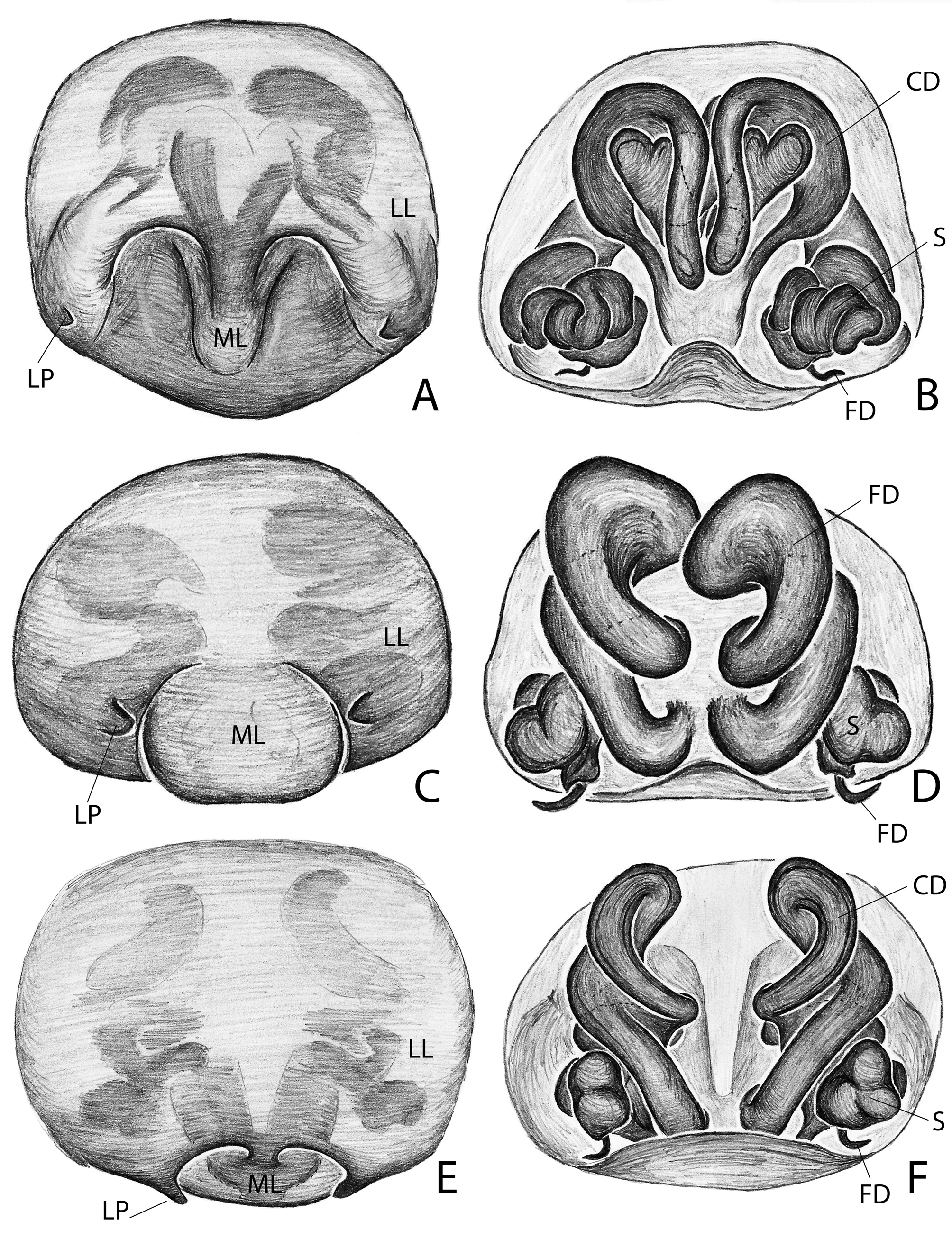
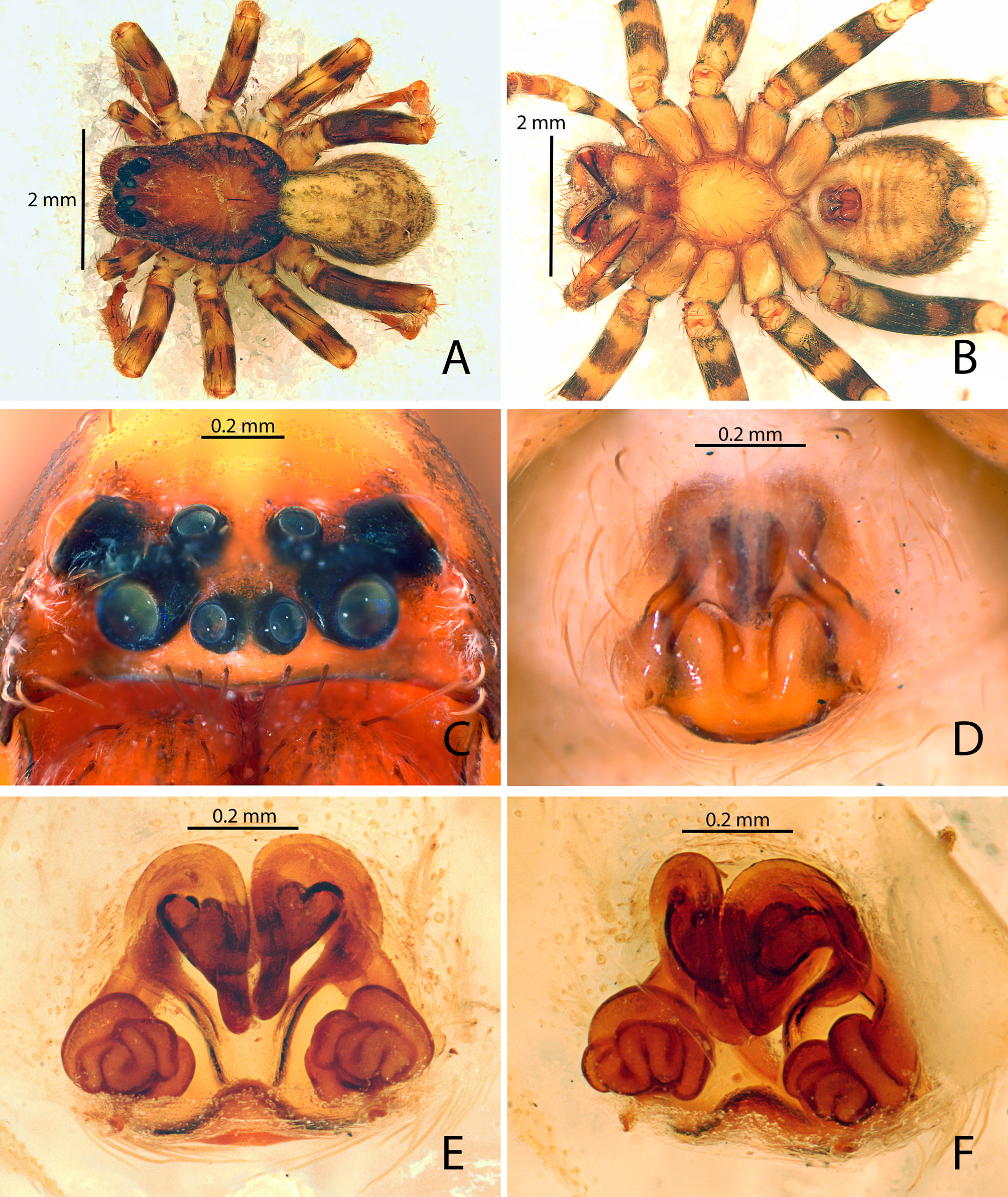
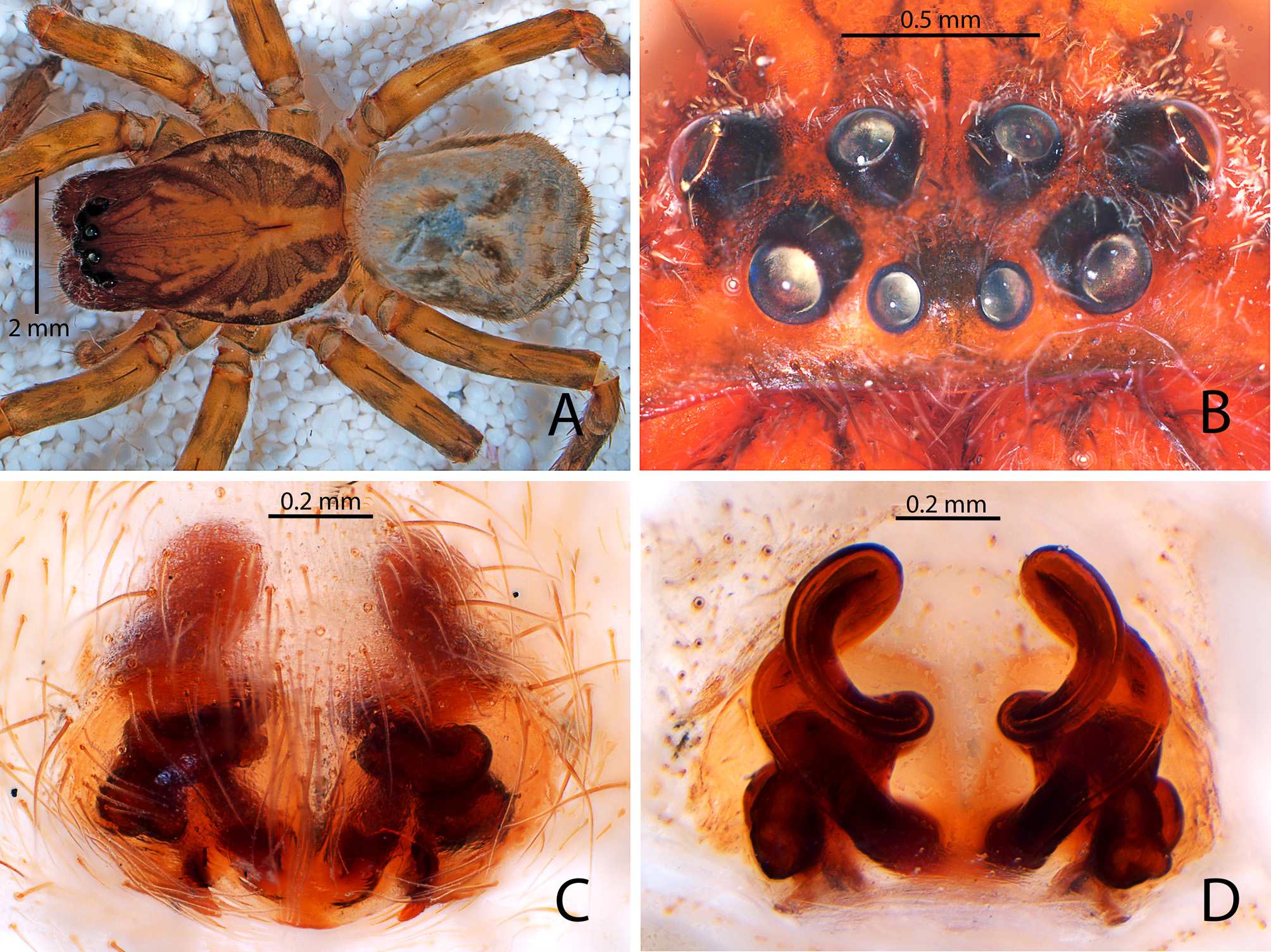
Diagnosis. Males of Campostichomma can be distinguished from those of other Udubidae by the palpal tibia length less than ½ length of cymbium ( Fig. 3A View FIGURE 3 ), the cymbium with a deep groove retrolaterally ( Fig. 4A, C View FIGURE 4 ) and the embolus elongate, threadlike, extending 360 ˚ around tegulum ( Figs 3D View FIGURE 3 , 4A View FIGURE 4 ). Females have a long, broad epigynal plate divided into medial and lateral lobes ( Figs 4D View FIGURE 4 , 8A–F View FIGURE8 ), lateral lobes with ( Fig. 8A, C, E View FIGURE8 ) or without teeth ( Fig. 4D View FIGURE 4 ), and internally the copulatory ducts have several loops, which can be used to identify the species of the genus ( Figs 4E View FIGURE 4 , 8B, D, F View FIGURE8 ).
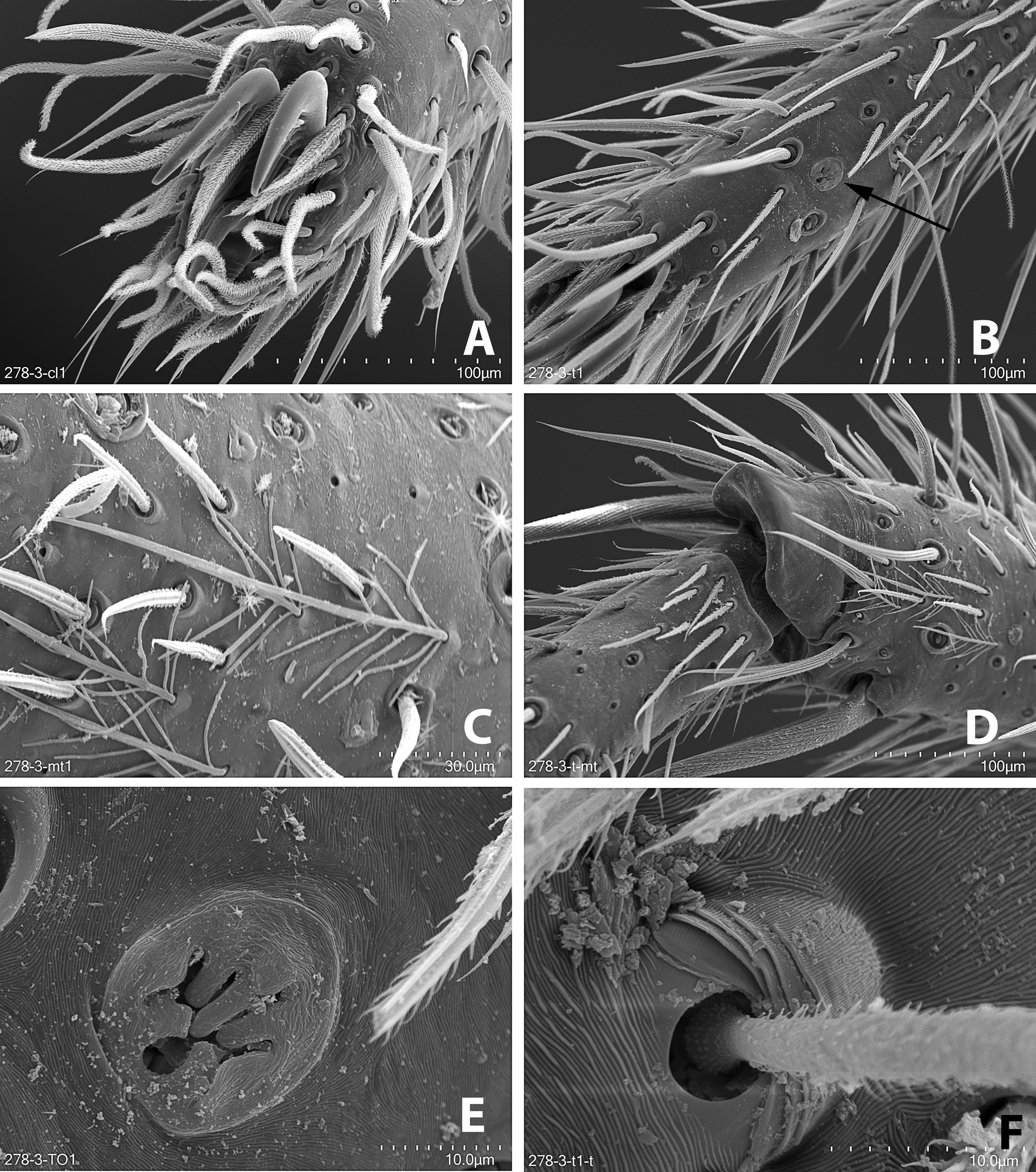
DesCription. Medium sized-spiders, total length 4.30–7.40; sexual dimorphism slight ( Fig. 2A–D View FIGURE2 ), males smaller with relatively longer legs. Carapace pear-shaped in dorsal view, domed; fovea linear, deep ( Fig. 2A, C View FIGURE2 ). Ocular area broad, ocular quadrangle trapezoidal, widest behind; eight eyes in 2 rows, anterior row slightly recurved, posterior row nearly straight ( Figs 2E–F View FIGURE2 , 5C View FIGURE5 , 6B View FIGURE 6 , 7B View FIGURE7 ). Clypeus low ( Figs 2E–F View FIGURE2 , 5C View FIGURE5 , 6B View FIGURE 6 , 7B View FIGURE7 ). Chelicerae stout with large boss, especially in female, pro- and retromargins of fang furrow with three and four stout teeth, respectively. Sternum broadly oval, bluntly curved or with small point posteriorly; labium with shallow basal notch; palpal coxae converging slightly anteriorly ( Figs 2B, D View FIGURE2 ), serrula in a single row along outer margin. Pedicel elongated and not fused ( Fig. 6C View FIGURE 6 ). Legs moderately long; leg formula 4123; integument finely wrinkled ( Griswold 1993: fig. 77), with plumose setae (sensu Lehtinen 1975) and feathery scales ( Fig. 1C View FIGURE 1 ; Griswold 1993: figs 75, 77); trochanters notched. Femora of legs and palpi with dorsal and lateral spines, patellae III and IV with lateral spines, tibiae I and II with 4 pairs of ventral spines and an additional proventral spine just behind apical pair, metatarsi I and II with 3 pairs of ventral spines, males usually with, females without, lateral spines on tibiae and metatarsi I and II; tibiae and metatarsi III and IV with dorsal, lateral, and ventral spines; palpal femora with dorsal and lateral spines, tibiae and tarsi of females with spines, tibiae and tarsi of males with or without spines, cymbium usually lacking spines. Males with tibial crack on legs I–IV ( Fig. 3E View FIGURE 3 ). Preening combs and scopulae absent. Superior tarsal claws pectinate, with inferior tarsal claws, I–II usually medium to large ( Fig. 1A View FIGURE 1 ; Griswold 1993: fig. 5), claw tufts absent ( Fig. 1A View FIGURE 1 ). Trichobothria on tibiae with basal dorsolateral group and pro- and retrolateral rows, retrolateral row extending to apex, metatarsi with dorsal, irregular row or dorsal row plus dorsobasal group, tarsi with dorsobasal row that divides into 2–3 rows apically; palpal tibiae with pro- and retrolateral rows, apparently absent from tarsi; trichobothrial base with transversely-ridged hood ( Fig. 1F View FIGURE 1 ; Griswold 1993: fig. 77). Tarsal organ median, capsulate, orifice oval to round or keyhole-shaped ( Fig. 1E View FIGURE 1 ; Griswold 1993: figs 76–77), with proximal seam. Male palpal tibia short. Cymbium with large, triangular petiole attached to alveolus; subtegulum cup-shaped, with anelli, prolateral margin with lobe that articulates with corresponding lobe on tegulum in unexpanded bulb ( Fig. 3B View FIGURE 3 ); tegulum C-shaped ( Fig. 4A View FIGURE 4 ), simple; firmly attached embolus (E) arising on retrolateral side of tegulum and tapering to apex; median apophysis (MA) concave, attached in middle of tegulum ( Fig. 3A View FIGURE 3 ); conductor (C) hyaline, originating anteriorly of tegulum, fan-shaped and not embracing apex of embolus ( Fig. 4A–C View FIGURE 4 ); fundus in subtegulum, reservoir and ejaculatory duct simple, without loops or switchbacks, spiralling around outer margin of tegulum. Abdomen oval, without scuta ( Fig. 2A–D View FIGURE2 ). Respiratory system comprising two anterior book lungs and a posterior tracheal spiracle. Epigynum divided by longitudinal epigynal folds into median lobe and lateral lobes; lateral lobes with or without teeth ( Figs 4D View FIGURE 4 , 8A, C, E View FIGURE8 ). Vulva with elongated copulatory duct, spermathecae with protuberances and small fertilization ducts ( Figs 4E View FIGURE 4 , 8B, D, F View FIGURE8 ). Cribellum absent; colulus a fleshy triangular lobe set with several setae. Six spinnerets, anterior (ALS) and posterior laterals (PLS) two-segmented, posterior medians (PMS) one-segmented; females with ALS having two major ampullate gland spigots and several piriform gland spigots, PMS with multiple minor ampullate, cylindrical and aciniform gland spigots; PLS with at least cylindrical and aciniform gland spigots.
Composition. Four species: C. manicatum Karsch, 1892 , C. alawala sp. nov., C. mudduk sp. nov. and C. harasbedda sp. nov.
Distribution. Endemic to Sri Lanka ( Fig. 14 View FIGURE14 ).
Natural history. The type specimens of C. manicatum were collected in the highlands of Sri Lanka. Collection data for the specimens collected by the Lund University Ceylon Expedition are detailed in Brinck et al. (1971). These data suggest that these are running spiders that favor moist, closed canopy forest or woodland. Most were collected in the highland above 1000 m, in forest, moist ravines or near tea plantations. C. alawala sp. nov. is unique in being from lower elevation, from jungle at only 200 m elevation. More detail is provided with each species description.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Campostichomma Karsch, 1892
| Polotow, Daniele & Griswold, Charles 2017 |
Campostichomma KArsch, 1892
| POlOtOw 2015: 151 |
| GrIswOld 1999: 59 |
| GrIswOld 1993: 7 |
| LEhtInEn 1967: 221 |