
Echinolittorina tuberculata ( Menke, 1828 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2184.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3606F-A54E-FFDA-FF26-FF27FA92FC02 |
|
treatment provided by |
Felipe |
|
scientific name |
Echinolittorina tuberculata ( Menke, 1828 ) |
| status |
|
Echinolittorina tuberculata ( Menke, 1828) View in CoL
( Figures 2E–G View FIGURE 2 , 31 View FIGURE 31 , 32 View FIGURE 32 , 33A–F View FIGURE 33 )
Trochus nodulosus Gmelin, 1791: 3582 View in CoL (“ Habitat in Oceano australi, et (minor) mari, Americam meridionalem alluente ”; in part; lectotype ( Clench & Abbott 1942) Chemnitz 1781: pl. 163, figs 1545, 1546, from Southern Ocean, i.e. Pacific Ocean, is E. pascua ( Rosewater, 1970) View in CoL or a member of the E. malaccana View in CoL group; Gmelin’s concept probably also included E. tuberculata View in CoL ; not Trochus nodulosus Solander View in CoL in Brander, 1766).
Littorina nodulosa —d’Orbigny, 1841: 205, pl. 14, figs 11–13, 14 (operculum) (not Gmelin, 1791).
Littorina (Tectarius) nodulosa —Arango, 1880: 160 (not Gmelin, 1791).
Litorina (Hamus) nodulosa View in CoL — Watson, 1886: 577–578 (in part, includes E. vermeiji View in CoL ; not Gmelin, 1791).
Tectarius nodulosus View in CoL — Tryon, 1887: 258, pl. 47, figs 59, 60, pl. 48, fig. 74 (in part, includes N. pyramidalis View in CoL , E. malaccana (Philippi, 1847) View in CoL , E. marquesensis Reid, 2007 View in CoL , Tectarius antonii ( Philippi, 1846 b) View in CoL ; not Gmelin, 1791).
Litorina tuberculata Menke, 1828: 25 View in CoL (no locality; neotype ( Bandel & Kadolsky 1982) USNM 749822, Pte des Châteaux, E tip Grande Terre, Guadeloupe, seen; Fig. 31A View FIGURE 31 herein).
Littorina (Tectus) tuberculata — Mörch, 1876: 133. Nevill, 1885: 156.
Nodilittorina tuberculata View in CoL — Abbott, 1954a: 134, pl. 19i. Abbott, 1954b: 451–456, fig. 55b, d (foot), f (operculum), h (faecal pellets), l (egg capsule), n (penis), 56a (map), 57b (radula). Coomans, 1958: 63, pl. 8. Abbott, 1968: 81–83, fig. Kaufman & Götting, 1970: 350, fig. 38. Borkowski, 1971: fig. 2 (egg capsule). Flores, 1973a: 10–11, pl. 1, figs 1–4. Bandel, 1974a: 100–101, 103, 108, figs 13 (shell), 16B and 17 (egg capsule), 18–22 and 39–41 (radula). Bandel, 1974b: 13, fig. 6C (faecal pellets). Bandel, 1975: 17, 18, pl. 2, figs 3, 5 (larval shell). Borkowski, 1975: 369–377, fig. 2B (radula). García-Talavera, 1983: 44–45 (in part, includes E. vermeiji View in CoL , E. helenae (E.A. Smith, 1890) View in CoL , E. granosa (Philippi, 1848) View in CoL , E. miliaris ( Quoy & Gaimard, 1833) View in CoL , Tectarius striatus View in CoL ). Bandel, 1984: fig. 14 (radula). Sterrer, 1986: 408, pl. 135. De Jong & Coomans, 1988: 20, pl. 32, fig. 84. Britton & Morton, 1989: 86, fig. 4- 5L. Díaz & Puyana, 1994: 126, pl. 34, fig. 409. Reid, 2002a: 259–281, figs 1D, 4 (pallial oviduct), 5A (radula).
Nodilittorina (Echinolittorina) tuberculata View in CoL — Habe, 1956: 96–99. Abbott, 1958: 35. Warmke & Abbott, 1961: 54, pl. 9r. Rehder, 1962: 123. Rosewater, 1970: 424, pl. 326, figs 10, 12. Abbott, 1974: 70, fig. 570. Rosewater, 1981: 33. H.E. Vokes & E.H. Vokes, 1983: 14 (in part, includes Tectarius antonii View in CoL ). Reid, 1989: 99, fig. 14j (radula).
Nodilittorina (Nodilittorina) tuberculata View in CoL — Bandel & Kadolsky, 1982: 32–34, figs 3B (operculum), 5 (radula), 10 (map), 13 (embryonic shell), 52–55 (shells and radulae).
Echinolittorina tuberculata View in CoL — Williams, Reid & Littlewood, 2003: 60–86. Williams & Reid, 2004: 2227–2251, fig. 6F (map).
Litorina thiarella Anton, 1838: 53 (no locality; holotype MTD 1641 View Materials a, not seen; Schniebs, 1995: fig. 4a, b).
Litorina (Tectarius) thiarella —Weinkauff, 1882: 45–46, pl. 5, figs 9, 11 (pl. 5 by Küster, 1856). Weinkauff, 1883: 225 (as Tectaria View in CoL ).
Littorina dilatata d’Orbigny, 1841: 207 View in CoL , pl. 14, figs 20–23 (la Havane [La Habana], Cuba; lectotype ( Bandel & Kadolsky 1982) + 14 paralectotypes BMNH 1954.10 .4.126, seen). Reeve, 1857: sp. 9, pl. 2, fig. 9.
Litorina dilatata — Philippi, 1846 b: 145, Litorina pl. 2, fig. 13.
Littorina (Tectus) dilatata View in CoL — Mörch, 1876: 133–134. Nevill, 1885: 156.
Littorina (Tectarius) dilatata View in CoL —Arango, 1880: 158–159.
Litorina (Tectarius) dilatata —Weinkauff, 1882: 47, 86, pl. 5, figs 17–19 (pl. 5 by Küster, 1856), pl. 12, figs 1, 4. Weinkauff, 1883: 225 (as Tectaria ).
Littorina (Nodilittorina) dilatata View in CoL —von Martens, 1897: 204.
Nodilittorina dilatata View in CoL — Bandel, 1974a: 100, 103, 108, figs 12 (shell), 17 (egg capsule), 18–22 and 48–49 (radula). Garcia & Luque, 1986: 37. Redfern, 2001: 28, pl. 14, fig. 114.
Nodilittorina (Granulilittorina) dilatata View in CoL — Rosewater, 1981: 33.
Nodilittorina (Nodilittorina) dilatata View in CoL — Bandel & Kadolsky, 1982: 29–31, figs 5 (radula), 10 (map), 42–44 (shells and radula).
Nodilittorina (Echinolittorina) dilatata View in CoL — Reid, 1989: 99, fig. 2g, 5g (head), 7j (penis), 8g (paraspermatozoa), 10h (pallial oviduct), 11l (egg capsule), 14k (radula).
Litorina trochiformis — Philippi, 1846 b: 143–144, Litorina View in CoL pl. 2, figs 12, 14, 15 (not Dillwyn, 1817 = a Pacific species).
Littorina (Tectarius) trochiformis —Arango, 1880: 160 (not Dillwyn, 1817).
Littorina (Nodilittorina) trochiformis —von Martens, 1897: 204 (not Dillwyn, 1817).
Tectarius trochiformis — Dall & Simpson, 1901: 431 (not Dillwyn, 1817). Johnson, 1934: 102 (not Dillwyn, 1817).
Echinella trochiformis — Lebour, 1945: 466–467, fig. 7 (egg capsule and larva) (not Dillwyn, 1817).
Echinella nodulosa var. matanzensis Richards, 1935: 257 , pl. 25, figs 26 (29 in error), 30, 31 (6.5 km northeast of Punta Sabanilla on Matanzas Bay , Matanzas Prov., Cuba, Quaternary ; holotype (fig. 30) Carnegie Museum, Pittsburgh, not seen; paratype ANSP 139736 About ANSP , not seen).
Tectarius tuberculatus (Wood) View in CoL — Clench & Abbott, 1942: 2–3, figs 1–4, 5 (operculum) (not Turbo tuberculatus Wood, 1828 View in CoL = Tectarius antonii View in CoL ). Lewis, 1960: 414–415, fig. 10A–C (egg capsule), D (veliger) (not Wood, 1828).
Echininus nodulosus View in CoL (‘L. Pfeiffer’)— Díaz & Puyana, 1994: 126, pl. 34, fig. 411 (not Gmelin, 1791; not ‘L. Pfeiffer, 1839 ’ = Tectarius antonii View in CoL ).
Taxonomic history: The confused identity of Trochus nodulosus Gmelin, 1791 View in CoL has been widely discussed ( Philippi 1846 b; Weinkauff 1882; Watson 1886; Clench & Abbott 1942; Abbott 1954b; Fischer 1967; Bandel & Kadolsky 1982; Reid & Williams 2004; Reid 2007). It is based on both a Pacific species (either E. pascua View in CoL or a member of the E. malaccana View in CoL group) and on E. tuberculata ( Reid 2007) View in CoL . The latter is suggested by Gmelin’s mention of a small form from seas of southern America: “ Habitat in Oceano australi, et (minor) mari, Americam meridionalem alluente, tuberculis minutis albis ” ( Gmelin, 1791; italics original). The name Trochus nodulosus View in CoL is, however, unavailable, being a junior primary homonym. Gmelin appears to have based his species entirely on the description and figures of Chemnitz (1781), and the locality Gmelin gave for the small form is therefore a misrepresentation of Chemnitz’ original ‘West Indies’. In this quotation from Gmelin, ‘minor’ is a descriptive term, not a new name (in this work, italic font is not used for Latin names; under ‘Habitat’ all text is in italics except for named localities). Furthermore, from the format of the entire work it is clear that “ tuberculis minutis albis ” is a description of the species Trochus nodulosus View in CoL as a whole, supplementary to the brief diagnosis, and does not apply just to the small form. Menke (1828) introduced the name Litorina tuberculata View in CoL for this small form, without description, but with the indication “ Trochus nodulosus View in CoL , minor, Gm.” This is interpreted as the introduction of a new species (not a replacement name), with a reference to the previously unnamed ‘small’ form that resembles Trochus nodulosus View in CoL except by its size and ‘American’ locality (this fulfils the requirement of ICZN 1999: Art. 12.2.1). The combination of Gmelin’s (1791) description and Chemnitz’ (1781) West Indian locality are sufficient to diagnose the present species. Since no original material associated with Chemnitz exists in ZMK (or in St Petersburg, Martynov 2002), Bandel & Kadolsky (1982) designated a neotype for Litorina tuberculata View in CoL in order to fix the concept of the species.
The synonymy does not include Turbo trochiformis Dillwyn, 1817 , which was proposed as a new name for Trochus nodulosus , because Dillwyn gave only ‘Southern Ocean’ (i.e. Pacific) as the type locality. At the end of the synonymy of Litorina trochiformis, Philippi (1846 b: 143) stated: ‘ var. minor est L. thiarella Anton’ (italics here indicate a widely spaced Roman font in the original). Bandel & Kadolsky (1982) interpreted this as the introduction of a new varietal name minor based on Anton’s species, but this was not Philippi’s intention. This is clear from the distinct typefaces and from comparison with the style of the text of Philippi’s (1846 b–1848) entire monograph of Litorina (no new varieties were named in plates 1 to 4; only in plate 5 were varieties named, and then using the characteristic widely spaced Roman typeface reserved for Latin names). The name ‘var. minor’ was also used, with a reference to L. thiarella , by Mörch (1876), but repeated use of this same term for other taxa, and the format of the work, demonstrate that this is an infrasubspecific category.
Shell shape in this species ranges from narrow with rounded nodules to broad and spinose with a wide pseudumbilicus. There has been a long tradition of recognizing these two extreme forms as distinct species. d’Orbigny (1841) used the name L. nodulosa for the former and described the latter as L. dilatata . This was followed by other nineteenth century authors, although various names were used for the narrower form: Litorina trochiformis by Philippi (1846 b), L. tuberculata by Mörch (1876; Nevill 1885) and Litorina thiarella by Weinkauff (1882, 1883). Tryon (1887) had a much broader species concept and united members of at least five species and three genera, from both the western Atlantic and Indo-Pacific, as Tectarius nodulosus , claiming that “it is impossible to distinguish them by any character”.
In their revision of the nodulose littorinids of the western Atlantic, Clench & Abbott (1942) considered the narrow and broad forms to be conspecific, but confusingly used the name ‘ Tectarius tuberculatus Wood’ (Wood’s species is Tectarius antonii , see below). This error was corrected by Abbott (1954b), who reinstated Menke’s name (for the first time since Mörch 1876 and Nevill 1885) in the combination N. tuberculata .
Detailed study of the egg capsules and radulae of this species revealed further variation. Bandel (1974a) sampled a broad, spinose form from the Bahamas and a narrower form with rounded nodules from Colombia and Curaçao. The radula of the former was typical of the group (as in Fig. 33A, B View FIGURE 33 ), but in the latter the rachidian was narrow and the major cusps of the lateral and inner marginal teeth enlarged (as in Fig. 33E, F View FIGURE 33 ). Furthermore the egg capsules were slightly different, the former with a broad peripheral band ( Fig. 32G View FIGURE 32 ) and the latter with the entire dome-shaped upper surface sculptured by rings ( Fig. 32H View FIGURE 32 ). Bandel therefore recognized these forms as distinct species, N. dilatata and N. tuberculata respectively. This was restated in the monograph of Bandel & Kadolsky (1982) and both names became established in the literature (e.g. Garcia & Luque 1986; Sterrer 1986; Britton & Morton 1989; Reid 1989; Redfern 2001). Rosewater (1981) even assigned the two taxa to different subgenera, Granulilittorina and Echinolittorina , on the basis of their contrasting radulae. However, in an assessment of variation of radulae and egg capsules in western Atlantic littorinids, Borkowski (1975) suggested that the two forms were extremes of the range of a single species. This was also the view of Reid (2002a), who tentatively combined the two, but noted that genetic data were required for confirmation. These data are provided here.
The sympatric species Tectarius antonii is superficially similar in appearance and the epithets tuberculatus and nodulosus have been applied to it also, resulting in some confusion. It was first described as Turbo tuberculatus Wood, 1828 (in the same year that Menke named Litorina tuberculata ) and used in the combination L. tuberculata (Wood) by d’Orbigny (1841) and Nina tuberculata (Wood) by Mörch (1876). However, this name is unavailable, because it is a primary homonym of Turbo tuberculatus Pennant, 1777 . Clench & Abbott (1942) mistakenly identified E. tuberculata (Menke) as Tectarius tuberculatus (Wood) , but this was subsequently corrected by Abbott (1954b). To add to the confusion between these two species, Pfeiffer (1839) incorrectly used the name Trochus nodulosus Gmelin, 1791 in the combination Litorina nodulosa for the species Tectarius antonii , and was subsequently erroneously credited as the author of a new name ‘ Litorina nodulosa L. Pfeiffer, 1839 ’, which became widely used for T. antonii (e.g. Philippi 1847; Clench & Abbott 1942; Abbott 1974). The nomenclature and complex synonymy of T. antonii have been clarified by Bandel & Kadolsky (1982). Occasional misidentifications have continued, however (H.E. Vokes & E.H. Vokes 1983; Díaz & Puyana 1994).
Diagnosis: Shell of medium to large size; sculptured with 2–3 rows of strong nodules. Penis with large glandular disc, three times size of adjacent mamilliform gland. Caribbean Sea , Bahamas, Florida, Bermuda. COI: GenBank AJ622995 View Materials , AJ622996 View Materials , FN298404 View Materials , FN298408 View Materials , FN298409 View Materials .
Material examined: 171 lots (including 31 penes, 17 sperm samples, 17 pallial oviducts, 29 radulae).
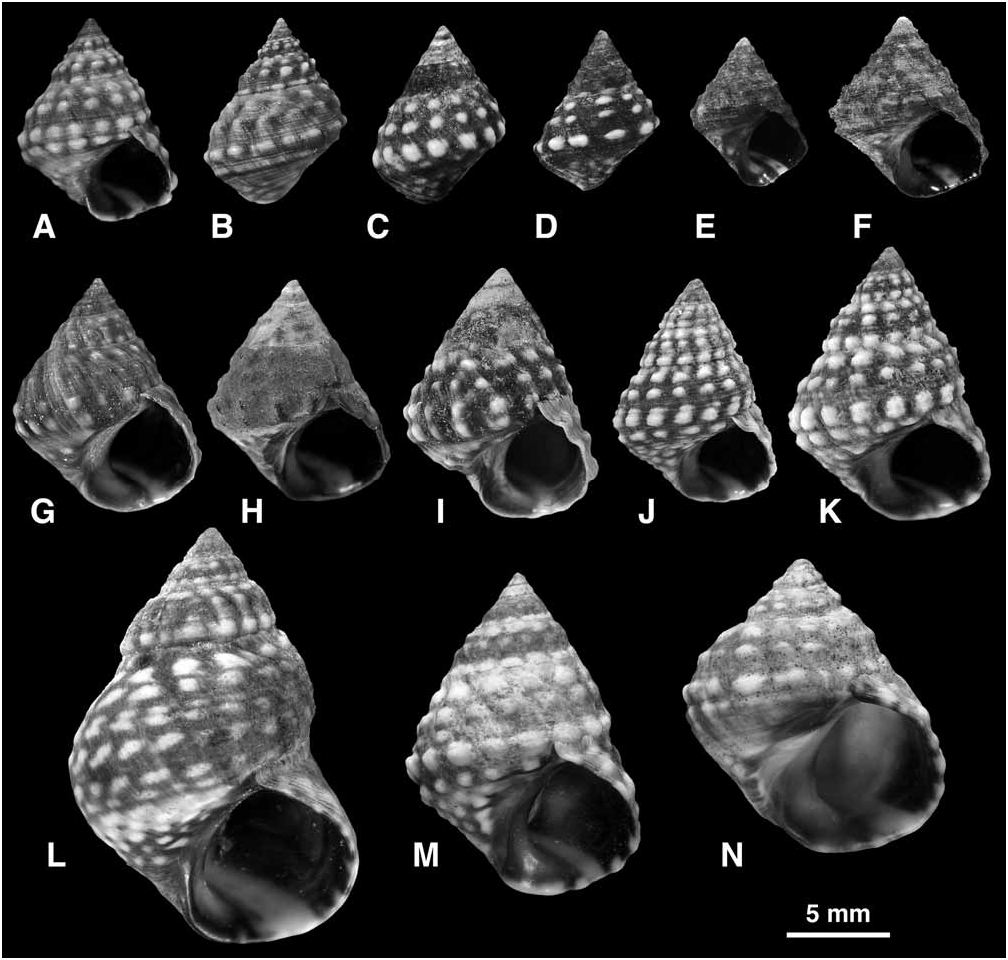
Shell ( Fig. 31 View FIGURE 31 ): Mature shell height 6.0– 22.9 mm. Shape turbinate to high turbinate (H/B = 1.14–1.75, SH = 1.49–2.20); spire whorls rounded; suture distinct; spire profile straight to slightly concave; periphery of last whorl rounded or slightly angled. Columella short, wide, hollowed and pinched at base, anterior lip may be expanded or slightly produced; narrow eroded parietal and columellar area; in broad shells the columellar area may be greatly widened and deepened, forming a funnel-shaped pseudumbilicus, outlined by a sharp flange continuous with the anterior lip ( Fig. 31J–L View FIGURE 31 ). Sculpture on spire whorls of 3 spiral rows of nodules (of which anterior may be partly covered by succeeding suture) overlain with fine spiral threads, nodules axially aligned; on last whorl 3 spiral rows of nodules remain prominent (at periphery, shoulder and a smaller row near suture); on base 3 (sometimes 2–4) rows of small nodules; narrow spiral threads cover entire surface together with fine spiral microstriae; in weakly sculptured shells the nodules at and above periphery remain small and rounded, and base may bear 4–6 non-nodulose cords ( Fig. 31A, O, P View FIGURE 31 ); in strongly sculptured shells nodules are represented by short, sharp spines (of which anterior face may be hollow) aligned on low axial ribs, and growth lines become minutely lamellose ( Fig. 31J, K View FIGURE 31 ); very large (gerontic) shells become more elongate (suture drops below periphery), primary nodules become smaller while a single row of secondary nodules develops between each, so towards end of last whorl sculpture consists of 10–12 subequal nodulose cords ( Fig. 31Q View FIGURE 31 ). Protoconch 0.28–0.31 mm diameter, 2.5–2.6 whorls. Ground colour grey brown, fading to blue grey (sometimes blackish when eroded), pale band on base, nodules white, cream or occasionally orange; aperture dark brown with pale band at base and sometimes a second band at shoulder; columella purple brown.
Animal: Head ( Fig. 32I View FIGURE 32 ) grey to black, sometimes an unpigmented stripe across snout; tentacle pale around eye and across base, with two longitudinal grey to black stripes up to full length of tentacle and a black dot at tip; sides of foot grey or black. Operculum ( Fig. 2E–G View FIGURE 2 ): opercular ratio 0.60–0.66 (lower in immature specimens, to 0.50; occasionally up to 0.76 in gerontic individuals), central part thickened. Penis ( Fig. 32A–E View FIGURE 32 ): filament about half total length of penis, with annular wrinkles for most of its length (so not clearly differentiated from wrinkled base), tapering or blade-shaped, sperm groove ends terminally; mamilliform gland about one-third size of large penial glandular disc, borne together on projection of base; penis unpigmented. Euspermatozoa 54–71 µm; paraspermatozoa ( Fig. 32J–L View FIGURE 32 ) containing 1–2 (rarely 4) parallelsided rod-pieces 16–28 µm (exceptionally to 38 µm), with rounded or truncated ends, usually projecting from cell, which is packed with round granules. Pallial oviduct ( Fig. 32F View FIGURE 32 ): copulatory bursa separates at posterior end of straight section and extends back between albumen and capsule glands; additional glandular material present in a swelling around egg groove at anterior end of straight section. Spawn ( Fig. 32G, H View FIGURE 32 ): an asymmetrically biconvex pelagic capsule 200–250 µm diameter by 100–130 µm high, with peripheral rim slightly overhanging base, dome-shaped upper side sculptured by 6–9 concentric rings or spiral whorls, containing single ovum 56–80 µm diameter ( Lebour 1945: Bermuda; Lewis 1960: Barbados; Borkowski 1971, 1975: Florida; Bandel 1974a: Bahamas, Colombia).
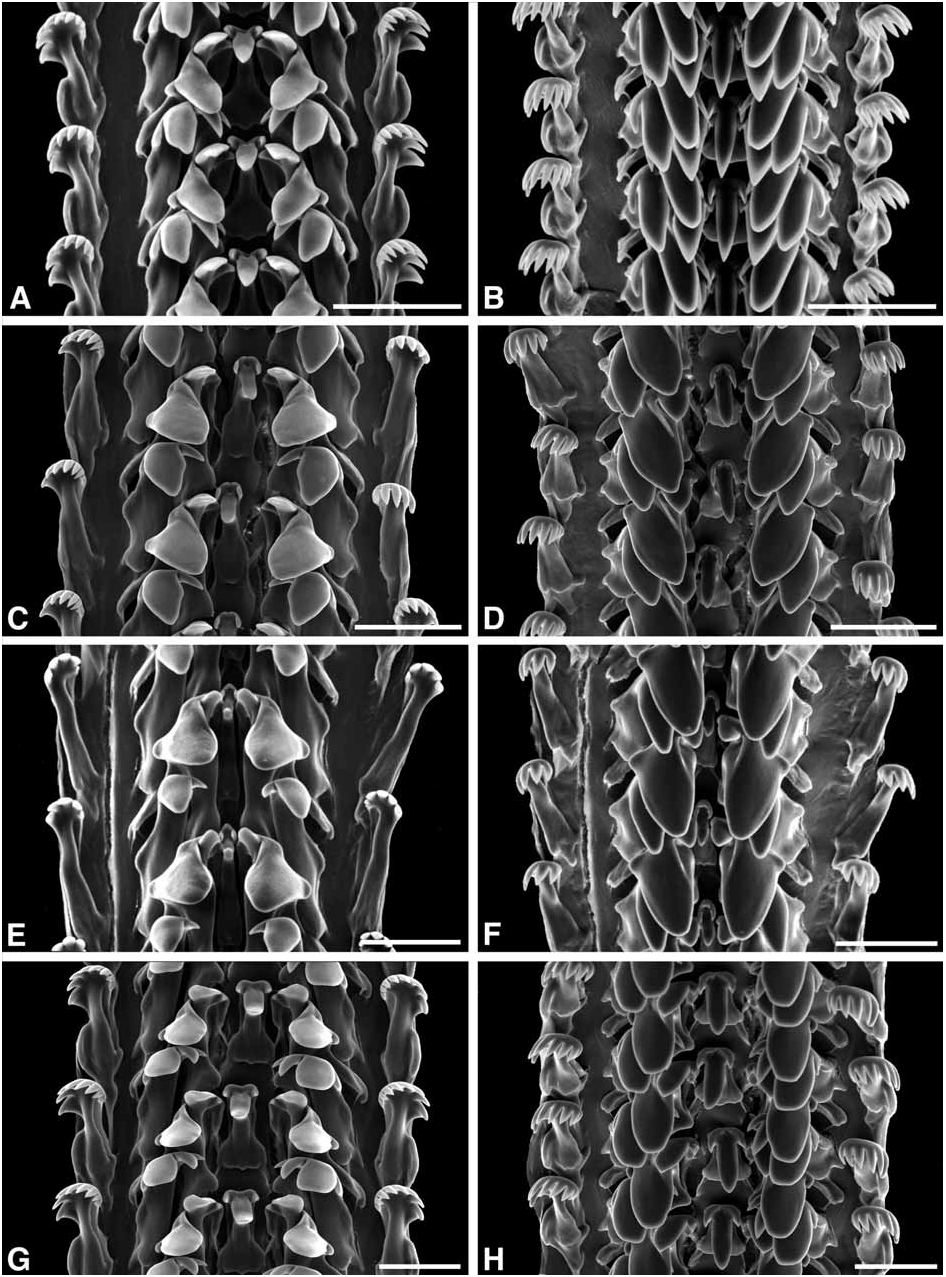
Radula ( Fig. 33A–F View FIGURE 33 ): Relative radula length 6.22–14.24 (longest radula 178 mm in shell 12.5 mm). Rachidian: length/width 2.00–4.40, sometimes narrow ( Fig. 32E, F View FIGURE 32 ); tip of major cusp pointed. Lateral and inner marginal: 4 cusps, tip of major cusp rounded; major cusp may be enlarged, innermost and outermost cusp of each tooth then absent ( Fig. 33C–F View FIGURE 33 ). Outer marginal: normal form with 7–8(9) cusps ( Fig. 33A View FIGURE 33 ), or narrowed to a rod with 5–6 cusps ( Fig. 33E, F View FIGURE 33 ).
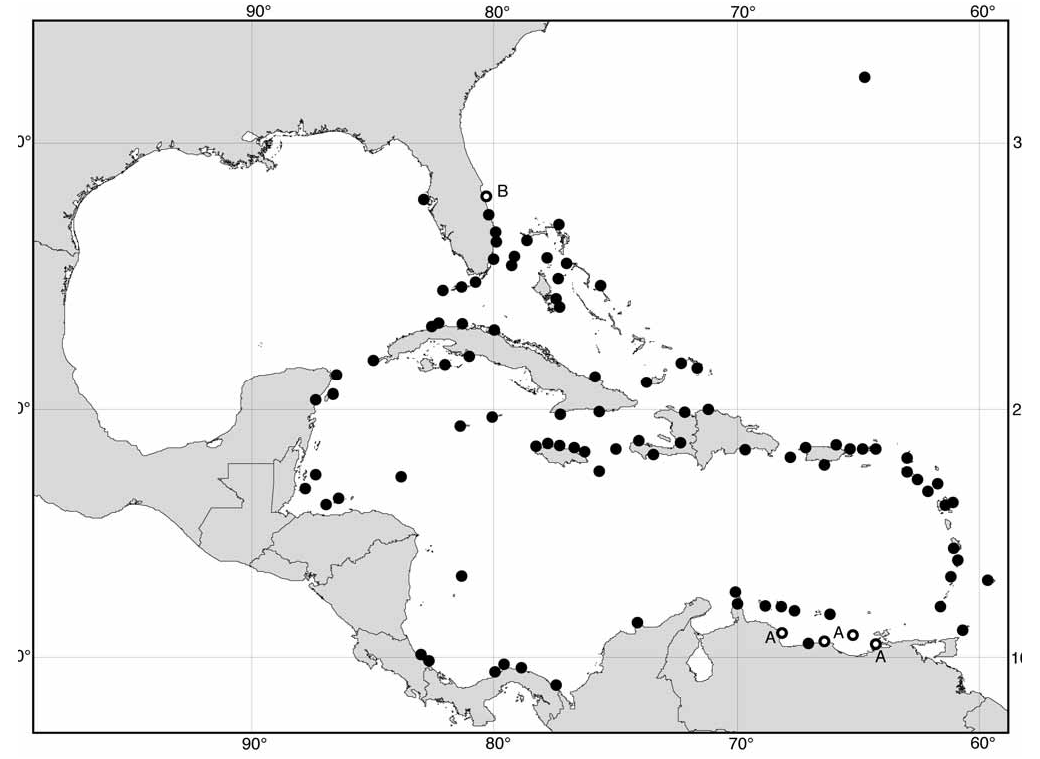
Range ( Fig. 34 View FIGURE 34 ): Throughout Caribbean Sea, Bahamas, S Florida, Bermuda. Range limits: Boca Ciega Key , Pinellas Co., Florida ( USNM 820455 About USNM ) ; Key West, Florida ( USNM 6409 About USNM ) ; Fort Pierce Inlet , St Lucie Co., Florida ( BMNH 20081042 ) ; Sebastian Inlet, Brevard Co., Florida (M. Krisberg, pers. comm.); Grape Bay , South Shore, Bermuda ( BMNH) ; Green Turtle Cay, Bahamas ( USNM 709139 About USNM ) ; Grand Caicos, Turks and Caicos Is ( USNM 391444 About USNM ) ; Pepillo Salcedo, Dominican Republic ( USNM 708544 About USNM ) ; Brewers Bay , St Thomas ( BMNH 20081062 ) ; Nenton Point, Antigua ( BMNH 20081040 ) ; Six Man’s Bay , Barbados ( BMNH) ; Grafton Bay , Tobago ( BMNH) ; Naiguata, E La Guaira, Venezuela ( RNHL) ; Aruba, Netherlands Antilles ( RNHL) ; Sta Marta, Colombia ( USNM 749824 About USNM ) ; Perme, NW Cape Tiburon , Panama ( USNM 664217 About USNM ) ; Playa Huertas, Portobelo, Panama ( BMNH) ; Cahuita, 40 km S Puerto Limón , Costa Rica ( BMNH) ; Isla de Providencia , Colombia ( USNM 103089 About USNM ) ; Swan Is, Honduras ( USNM 83647 About USNM ) ; Roatan I., Honduras ( BMNH 1961736 ) ; South Water Cay , Belize ( BMNH 20081043 ) ; 5 km S Tulum , Quintana Roo, Mexico ( BMNH 20081041 ) ; Isla Mujeres, Quintana Roo, Mexico ( USNM 662239 About USNM ) ; Morant Reef, Jamaica ( BMNH) ; Fort Charlotte , Lucea, Jamaica ( BMNH) ; Georgetown, Grand Cayman, Cayman Is ( BMNH 1961737 ) ; Gibara, Cuba ( IRSNB) ; Playa Giron, S Cuba ( BMNH 20081060 ) ; Vedado, Habana Prov., Cuba ( IRSNB) .
This species is common on all the islands of the Caribbean Sea, the Bahamas and Bermuda. Records on the American mainland are restricted to relatively oceanic sites, including La Guaira, Venezuela; Sta Marta, Colombia; and the NE extremity of the Yucatan Peninsula. Flores (1973b) found it to be more common on island than continental coasts in Venezuela. In Panama and Costa Rica it is scarce. The sole record from the west coast of Florida (Boca Ciega Key) requires confirmation, because this oceanic species would not be expected in the Gulf of Mexico.
Habitat: In the littoral fringe on coral limestone, beachrock and concrete; on sheltered and exposed shores; mainly in oceanic settings with clear water.
Among the western Atlantic Echinolittorina species this occupies the highest levels on the shore ( Flores 1973b; Vermeij 1973a; Bandel 1974a; Britton & Morton 1989; Lang et al. 1998; Brattström 1999). In Panama it extends up to the same level as Cenchritis muricatus , the most landward of all the littorinids on western Atlantic shores (Brattström 1986), but is usually lower. At Santa Marta, Colombia, it prefers horizontal ledges in the littoral fringe on shale cliffs, and may remain dry and attached only by a mucous film for long periods ( Bandel 1974a; Brattström 1980; Bandel & Wedler 1987). In the Bahamas it occupies the littoral fringe on beachrock and limestone, extending from the grey zone to the black zone below ( Brattström 1992, 1999). On limestone in the Florida Keys it extends from the edge of the white zone, through the grey zone and is most frequent in the black zone, but occurs no lower (T.A. Stephenson & A. Stephenson 1950). Some authors have recorded that E. tuberculata has a wider vertical range than other congeners ( Bandel & Kadolsky 1982; Lang et al. 1998); for example, it extends from the upper littoral fringe down to a zone of mosslike algae and to the top of the eulittoral in Jamaica ( Vermeij 1973b), and on limestone platforms in the Bahamas is found (in descending order) in the white, grey, black and upper yellow zones, but especially in the grey zone ( Newell et al. 1959). Smaller animals occupy lower levels in this range ( Lewis 1960; Vermeij 1972b; Bandel 1974a) and are more often found in crevices ( Lang et al. 1998). Yipp & Carefoot (1988) reported that the zonation level expanded downwards after removal of other littorinids, suggesting possible competitive effects. This is the most abundant of the littoral fringe species in Jamaica, where it occurs on both high and low energy shores ( Minton & Gochfeld 2001). Other authors have reported that E. tuberculata is rare or absent on sheltered shores, as in Colombia ( Bandel 1974a) and Bermuda ( Thomas 1985).
As well as occupying pits and crevices, E. tuberculata is found submerged in warm, saline pools in the littoral fringe ( Kristensen 1965; Vermeij 1973b; Bandel & Wedler 1987; Lang et al. 1998). This may account for its salinity tolerance of 34–40 ppt, which is higher than that of other sympatric littorinids ( Flores 1973b). Bandel (1974a) observed that juveniles crawled out of pools that were diluted by rain, whereas Lang et al. (1998) found snails in pools with nearly fresh water as well as in hypersaline pools, and suggested that this was for the purpose of rehydration. Coomans (1962) observed that animals died in less than a week if kept submerged in seawater, whereas Yipp & Carefoot (1988) reported survival for more than two weeks. Survival is greater in air, up to 32 days ( Yipp & Carefoot 1988; Britton 1992) or even 12 months ( Mattox 1949; although there may have been confusion with the more resistant Tectarius antonii, Abbott 1954b ).
The heat coma temperature is 45.4°C ( Britton 1992; McMahon 2001), while 50% survived for an hour at 49–50°C ( Fraenkel 1968). Surprisingly, this tolerance is lower than that of smooth-shelled congeners that occupy lower levels, perhaps because nodulosity encourages convective heat loss, reducing the tissue temperatures experienced ( Vermeij 1973a; Britton 1992). A detailed study by Lang et al. (1998) in Jamaica reported thermoregulation both above and below ambient temperature (body above ambient temperature when substrate less than 29°C, below when ambient temperature above 33°C), but noted that even in July the substratum temperature was always below the heat coma temperature.
The diet includes a predominance of thick-walled unicellular algae ( Yipp & Carefoot 1988) and the growth rate is slow ( Lewis et al. 1969; Borkowski 1974). McLean (1967) estimated erosion of beachrock by grazing at 1.2 g /year (the highest among the Echinolittorina species tested), and Dobson-Moore & Britton (2001) measured the rate of bioerosion on a limestone platform in Jamaica. It may forage mainly at night ( Lewis 1960). Predation on E. tuberculata by lampyrid glow-worms has been reported in Jamaica ( McDermott 1953). Abbott (1954b) mentioned occurrence of parasitic cercariae in animals from Cuba, but this is remarkably rare in Echinolittorina species ( Reid 2007) .
The breeding season extends through most of the year. In Florida, Borkowski (1971) found some males mature all year except February, while peak female maturity was reached between April and November. Breeding occurs throughout the year in Barbados ( Lewis 1960) and in Colombia ( Bandel 1974a), whereas at the northern extremity of the range in Bermuda capsules were reported in July and August ( Lebour 1945). Veligers hatch from the pelagic capsules after three ( Lewis 1960) or four days ( Bandel 1974a).
Remarks: A number of littorinine genera and species groups, without close phylogenetic relationship, show similarly nodulose shells ( Reid 2002 a, b, 2007; Williams et al. 2003; Williams & Reid 2004). These are usually the species that occupy the highest supratidal levels on the shore, and this shell form is interpreted as an adaptation to reduce tissue temperature by convective heat loss while the animal is attached only by a mucous film or ‘holdfast’ ( Vermeij 1973a). This, together with behavioural means, allows a degree of thermoregulation ( Lang et al. 1998). As an additional probable adaptation to the high-level habitat, the operculum is more tightly coiled (opercular ration 0.60–0.66) than that of any other Echinolittorina ; this serves the dual function of thickening the opercular material and fitting it to a more nearly circular aperture ( Bandel & Kadolsky 1982) reducing both the rate and the area for water loss.
The shell is superficially similar to that of a number of other nodulose high-shore littorinids, including the Australian Nodilittorina pyramidalis ( Reid & Williams 2004) , Tectarius antonii and Cenchritis muricatus in the Caribbean Sea, members of the E. natalensis and E. malaccana groups in the Indo-West Pacific ( Reid 2004), E. galapagiensis ( Stearns, 1892) in the eastern Pacific ( Reid 2002b), and E. miliaris , E. helenae and E. granosa in the eastern Atlantic ( Rosewater 1981; Bandel & Kadolsky 1982). Differences can be subtle, but the three rows of rounded nodules on the last whorl and relatively broad columella are usually diagnostic of E. tuberculata . In the western Atlantic the only other nodulose Echinolittorina species is E. vermeiji from islands off Brazil, which has a more strongly angled periphery and usually four rows of white nodules on a darker shell ( Fig. 35 View FIGURE 35 ). Confusion is unlikely to arise since none of the nodulose congeners is sympatric with E. tuberculata . However, Tectarius antonii is not only sympatric but also frequently syntopic with E. tuberculata , and there has been a history of confusion between the two (see Taxonomic History above). The shell of T. antonii is broader, the last whorl sculptured by two rows of large pointed tubercles or stout spines, the aperture more circular, the columella narrower and not anteriorly expanded ( Bandel & Kadolsky 1982). Although most shells of these two species are readily separated after some experience, some eroded shells may be ambiguous. The living animals can then be most easily separated by the thicker and more tightly wound, multispiral, operculum of T. antonii (opercular ratio = 0.78–0.81, cf. 0.61–0.66 (exceptionally 0.76) in E. tuberculata ). There are other anatomical differences: in T. antonii the penis bears 3–4 penial glands and a mass of papillae, and the capsule gland of the pallial oviduct occupies half of the total length of the straight section ( Reid 1989). Confusion with Cenchritis muricatus is unlikely; this shell is larger and broader, greyish white, with white aperture and four rows of granules on each spire whorl, found on wood as well as rock in the high littoral fringe, and the penis is simple, lacking obvious glands.
Intraspecific variation in the tooth form of the radula is so great that the form of normal Echinolittorina type ( Fig. 33A, B View FIGURE 33 ) was considered to belong to a different species (‘ N. dilatata ’) from that characterized by a narrowed rachidian ( Fig. 33E, F View FIGURE 33 ) ( Bandel 1974a; Bandel & Kadolsky 1982). Adding weight to this interpretation, there appeared to be correlated differences in the shell and spawn. The shell of N. dilatata was described as broader, having the nodules more sharply pointed, and the columella more dilated, sometimes with a pseudumbilicus ( Bandel & Kadolsky 1982; e.g. Fig. 31K View FIGURE 31 ), compared with the more elongate, imperforate shell of N. tuberculata with rounded nodules (e.g. Fig. 31A View FIGURE 31 ). The egg capsule of N. dilatata was characterized by six spiral rings and a broad peripheral rim ( Fig. 32G View FIGURE 32 ), as opposed to six to seven or more rings on a hemispherical cupola in N. tuberculata ( Fig. 32H View FIGURE 32 ) ( Bandel 1974a). Finally N. dilatata was considered to be distributed in Florida, the Bahamas and the Greater Antilles, while N. tuberculata was present in the Caribbean Sea ( Bandel & Kadolsky 1982). However, in an assessment of variability in western Atlantic littorinids, Borkowski (1975) dismissed this morphological evidence, considering the two ‘species’ to be extremes of a continuum. The variation in radular tooth dimensions of N. tuberculata was no greater than that in other species, although he apparently did not find such extreme ‘narrowed’ radulae as that reported by Bandel (1974a). Both forms of egg capsule could be found at a locality in Florida ( Borkowski 1971, 1975). Geographical distribution of the two potential species remained poorly known, but Reid (1989) found radular differences between sympatric individuals, and therefore accepted the two species as valid. However, evidence began to accumulate of the remarkable plasticity of radular form in some other littorinid genera ( Padilla 1998; Reid & Mak 1999). This, and the absence of differences in reproductive anatomy led Reid (2002a) to propose that N. dilatata and N. tuberculata were conspecific.
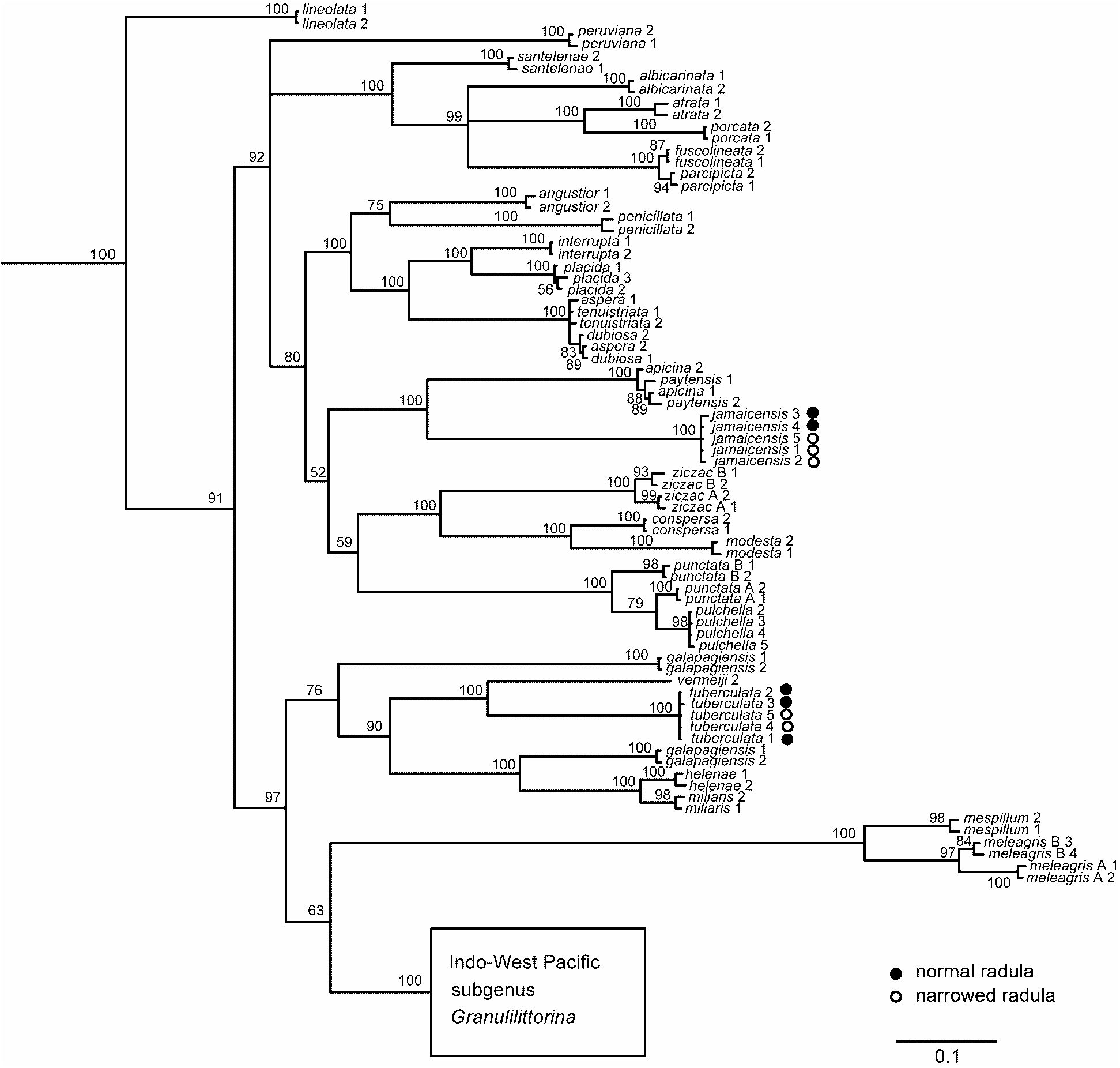
Sequence data analysed by Williams & Reid (2004) came only from two specimens with dilatata - type radulae, from Quintana Roo and Jamaica. COI sequence data from two individuals with narrowed tuberculata - type radulae from Barbados, and another with dilatata - type radula from Bermuda, have now been added, and all five cluster tightly together ( Fig. 37 View FIGURE 37 ; average K2P distance = 0.13%, range 0–0.25%), supporting conspecificity.
It remains to be determined what controls the intraspecific variation in radular form of E. tuberculata . In other littorinids there is direct ( Padilla 1998; Andrade & Solferini 2006a) or indirect ( Reid & Mak 1999) evidence of an ecophenotypic effect of substrate upon which the animals graze. In the present study 29 radulae have been examined (by light or electron microscopy). The normal dilatata - type is the more common (66%), found throughout the geographical range (Florida, Bermuda, Bahamas, Jamaica, Lesser Antilles, Panama, Mexico) and recorded from both limestone and concrete substrates. The narrowed tuberculata - type is present in the Bahamas, southern Cuba and Barbados, to which may be added Colombia and Curaçao (from metamorphic shale and beachrock; Bandel 1974a), Puerto Rico and the Lesser Antilles ( Bandel & Kadolsky 1982). There is therefore no clear geographical pattern, and no obvious correlation with substrate. The present study has revealed no connection with size or sex, and the type of radula cannot be reliably predicted from the shape or sculpture of the shell. The relative radular length is considerable, up to 14 times the length of the shell, almost as great as the 15 times reported in E. aspera ( Reid 2002b) , but again there is no correlation with radular type. The explanation for this variation remains a mystery, and further study is required.
Variation in the form of the egg capsule has been mentioned above. The type with a broad peripheral rim and six spiral rings ( Fig. 32G View FIGURE 32 ) has been recorded from Bermuda ( Lebour 1945) and the Bahamas ( Bandel 1974a), and the domed type with seven to nine rings ( Fig. 32H View FIGURE 32 ) from Barbados ( Lewis 1960) and Colombia ( Bandel 1974a). Both types were found in Florida ( Borkowski 1971, 1975). Comparable variation is seen in the spawn of E. angustior ( Borkowski 1971, 1975), E. natalensis (Philippi, 1847) ( Reid 2007) and E. meleagris ( Fig. 4K–M View FIGURE 4 ). Lewis (1960) reported that smaller capsules were sometimes produced in long strings; these are believed to be abnormal, probably released prematurely in the laboratory.
Shell variation in this species shows some regional trends. Shells from Bermuda attain the largest size ( Fig. 31M, Q View FIGURE 31 ). Those from Bermuda, the Bahamas and Cuba exhibit the strongest development of spines, and only here are found the uncommon extreme forms with relatively low spire, dilated columella and pseudumbilicus ( Fig. 31J–L View FIGURE 31 ). The low spire and pseudumbilicus are the consequence of an increased rate of displacement of the apertural generating curve from the axis of coiling; if the displacement rate is low an eroded parietal area results. The eroded parietal area and pseudumbilicus are therefore mutually exclusive ( Reid 2002a). In the southern half of the range, from Mexico to Jamaica, the Lesser Antilles and Venezuela, the shells tend to be slightly narrower, with smaller and more rounded nodules instead of sharp spines, and lack a pseudumbilicus ( Fig. 31A–D, H, N–P View FIGURE 31 ). This geographical pattern contributed to the decision of Bandel & Kadolsky (1982) to separate the northern form as N. dilatata . However, there are many exceptions and intermediates, and variation within local populations is often as high as between the most distant localities. In nodulose Indo-Pacific species of Echinolittorina it has been suggested that occurrence on a limestone substrate is correlated with strong shell sculpture, perhaps through an ecophenotypic effect ( Reid 2007). Whether a similar correlation exists in E. tuberculata is not known. It cannot, however, explain the geographical trend, because through most of its range the species is found on limestone substrates. Abbott (1954b) reported sexual dimorphism in Puerto Rico, females being slightly larger.
In the pioneering ecological surveys of the Florida Keys by T.A. Stephenson & A. Stephenson (1950) E. tuberculata was one of the 20 most common species recorded. However, in 1999 it was not encountered during a resurvey of five of the same sites by Smith, Purcell & Barimo (2007). The authors suggested that this could have been due to the activities of shell collectors, but this seems hardly credible. An alternative explanation is that the species could be sensitive to eutrophication, consistent with the report of increased abundance of algal indicator species. The species may have become less common, but has evidently not disappeared from the Keys; M. Krisberg (pers. comm.) has 14 records from the Keys since 2000.




| IRSNB |
Institut Royal des Sciences Naturelles de Belgique |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Echinolittorina tuberculata ( Menke, 1828 )
| Reid, David G. 2009 |
Echinolittorina tuberculata
| Williams, S. T. & Reid, D. G. 2004: 2227 |
| Williams, S. T. & Reid, D. G. & Littlewood, D. T. J. 2003: 60 |
Echininus nodulosus
| Diaz, J. M. & Puyana, M. 1994: 126 |
Nodilittorina (Nodilittorina) tuberculata
| Bandel, K. & Kadolsky, D. 1982: 32 |
Nodilittorina (Nodilittorina) dilatata
| Bandel, K. & Kadolsky, D. 1982: 29 |
Nodilittorina (Granulilittorina) dilatata
| Rosewater, J. 1981: 33 |
Nodilittorina dilatata
| Redfern, C. 2001: 28 |
| Garcia, M. T. & Luque, A. A. 1986: 37 |
| Bandel, K. 1974: 100 |
Nodilittorina (Echinolittorina) tuberculata
| Reid, D. G. 1989: 99 |
| Vokes, H. E. & Vokes, E. H. 1983: 14 |
| Rosewater, J. 1981: 33 |
| Abbott, R. T. 1974: 70 |
| Rosewater, J. 1970: 424 |
| Rehder, H. A. 1962: 123 |
| Warmke, G. L. & Abbott, R. T. 1961: 54 |
| Abbott, R. T. 1958: 35 |
| Habe, T. 1956: 96 |
Nodilittorina tuberculata
| Reid, D. G. 2002: 259 |
| Diaz, J. M. & Puyana, M. 1994: 126 |
| Britton, J. C. & Morton, B. 1989: 86 |
| De Jong, K. M. & Coomans, H. E. 1988: 20 |
| Sterrer, W. 1986: 408 |
| Garcia-Talavera, F. 1983: 44 |
| Bandel, K. 1975: 17 |
| Borkowski, T. V. 1975: 369 |
| Bandel, K. 1974: 100 |
| Bandel, K. 1974: 13 |
| Flores, C. 1973: 10 |
| Abbott, R. T. 1968: 81 |
| Coomans, H. E. 1958: 63 |
| Abbott, R. T. 1954: 134 |
| Abbott, R. T. 1954: 451 |
Echinella trochiformis
| Lebour, M. V. 1945: 466 |
Tectarius tuberculatus
| Lewis, J. B. 1960: 414 |
| Clench, W. J. & Abbott, R. T. 1942: 2 |
Echinella nodulosa var. matanzensis
| Richards, H. G. 1935: 257 |
Tectarius trochiformis
| Johnson, C. W. 1934: 102 |
| Dall, W. H. & Simpson, C. T. 1901: 431 |
Littorina (Nodilittorina) dilatata
| Martens, E. von 1897: 204 |
Littorina (Nodilittorina) trochiformis
| Martens, E. von 1897: 204 |
Tectarius nodulosus
| Tryon, G. W. 1887: 258 |
Litorina (Hamus) nodulosa
| Watson, R. B. 1886: 577 |
Litorina (Tectarius) thiarella
| Weinkauff, H. C. 1883: 225 |
Littorina (Tectus) tuberculata
| Nevill, G. 1885: 156 |
| Morch, O. A. L. 1876: 133 |
Littorina (Tectus) dilatata
| Nevill, G. 1885: 156 |
| Morch, O. A. L. 1876: 133 |
Litorina tuberculata
| Menke, K. T. 1828: 25 |
Trochus nodulosus
| Gmelin, J. F. 1791: 3582 |