
Chaetozone elakata, Lavesque, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4312.3.10 |
|
publication LSID |
lsid:zoobank.org:pub:Bb8621E2-A2Aa-48A8-9Bc5-2E8384Da88Ef |
|
DOI |
https://doi.org/10.5281/zenodo.6049829 |
|
persistent identifier |
https://treatment.plazi.org/id/8E899916-796B-473E-B7EE-C1A51E771D4B |
|
taxon LSID |
lsid:zoobank.org:act:8E899916-796B-473E-B7EE-C1A51E771D4B |
|
treatment provided by |
Plazi |
|
scientific name |
Chaetozone elakata |
| status |
sp. nov. |
Chaetozone elakata View in CoL new species
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3
Material examined. Northeast Atlantic Ocean, offshore France, Bay of Biscay. R/ V Côtes de la Manche Sta. JERICO-8, 45°38′55″N, 01°45′47″W, coll. 28 October 2016, 0.25m ² Hamon grab, 62 m, holotype ( MNHN-IA- TYPE 1800 ), 3 paratypes (MNHN-IA-TYPE 1801), 2 Paratypes, SEM (MNHN-IA-TYPE 1802) and 7 paratypes ( MCZ 143234 About MCZ ). GoogleMaps
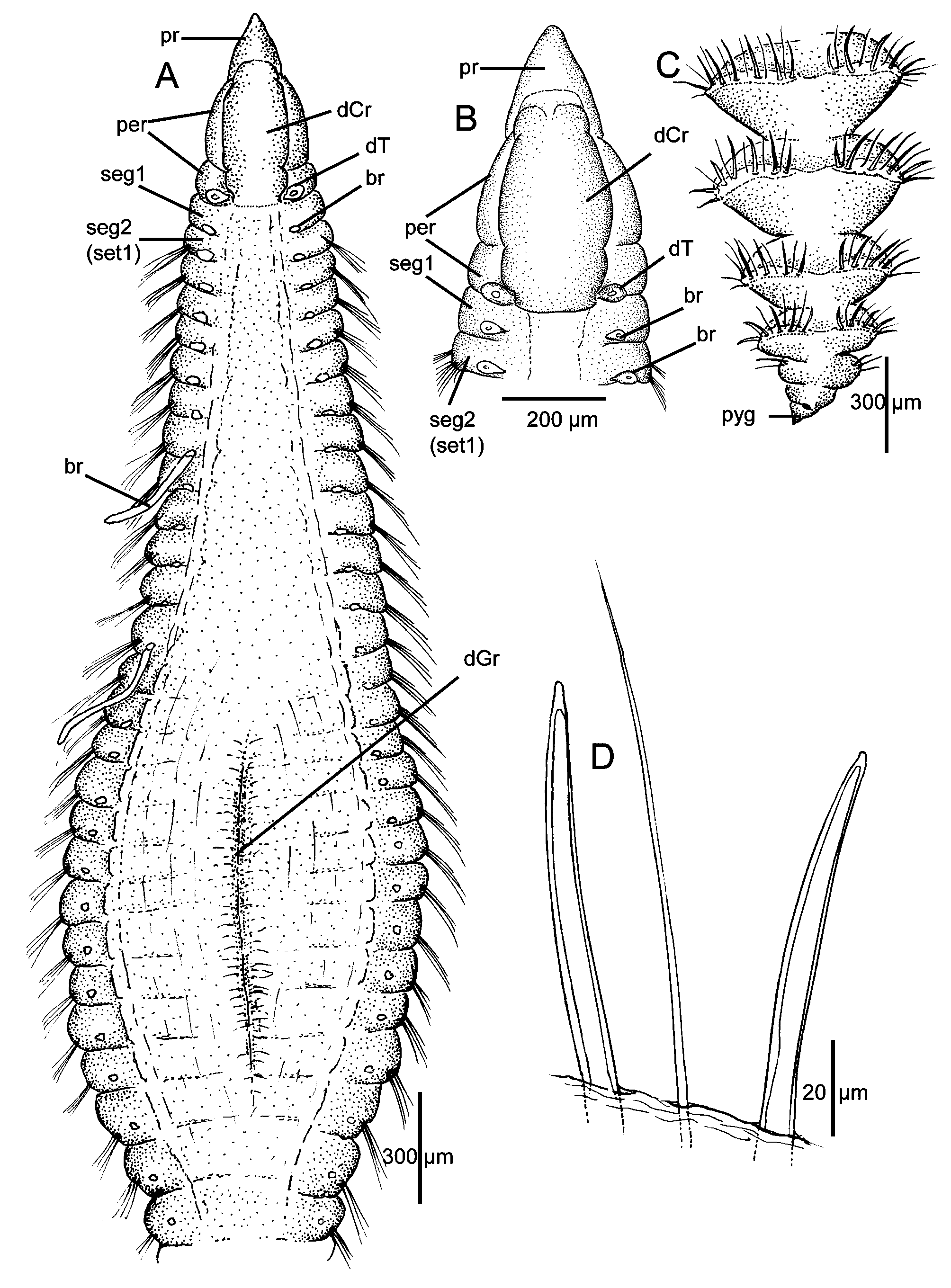
Description. A moderate-sized species with elongate body and conspicuous bulge in middle of thoracic region imparting distinct fusiform shape to anterior end ( Figs. 1 View FIGURE 1 A, 3A, D). Anterior segments crowded, about four times as wide as long ( Fig. 1 View FIGURE 1 A), then becoming longer, but still about twice as wide as long in middle body segments ( Fig. 3 View FIGURE 3 A, D). Posterior segments narrowing, with parapodia becoming modified, with deep grooves between individual segments ( Figs. 1 View FIGURE 1 C, 2F) and parapodia elevated with thin membranes bearing spreading fascicles of spines and alternating capillaries producing a prominent armature ( Fig. 2 View FIGURE 2 B–C).
Holotype (MNHN-IA-TYPE 1800) complete, 45.4 mm long, 0.64 mm wide anterior to bulge, 1.01 mm across bulge, 0.65 mm posterior to bulge, with 111 setigerous segments; complete paratype an ovigerous female (MCZ 143234) 33 mm long, 0.4 mm wide anterior to bulge, 0.65 across bulge, 0.52 mm posterior to bulge, with 117 setigerous segments. Other complete paratypes with 92–104 setigerous segments. Most specimens with distinct dorsal groove, in expanded segments ( Figs. 1 View FIGURE 1 A, 3B), absent in some specimens ( Fig. 3 View FIGURE 3 A); weak depression present on individual segments along most of venter; absent in posterior segments. Color in alcohol light tan with no apparent pigment on body; numerous segmental subdermal glands visible under a coverslip at about 63x magnification; these stain with Methyl Green imparting distinct patterns (see below).
Prostomium triangular, narrowing to rounded tip ( Fig. 1 View FIGURE 1 A–B); nuchal organs reduced to thin slits at posterior margin of prostomium, best seen with SEM ( Fig. 2 View FIGURE 2 A); eyespots absent. Peristomium relatively narrow, divided into two distinct annular rings ( Fig. 1 View FIGURE 1 A–B); first long, relatively smooth but with 2–3 weak lateral grooves best seen in SEM ( Fig. 2 View FIGURE 2 A), second ring narrow, similar in size and appearance to following achaetous segment. Entire peristomium surmounted by a relatively smooth, elongate dorsal crest extending posteriorly to anterior border of achaetous segment 1 ( Figs. 1 View FIGURE 1 A–B, 2A); dorsal tentacles arising from posterior margin of second peristomial ring ( Fig. 1 View FIGURE 1 A–B). First segment achaetous with first pair of branchiae arising from posterior margin; second segment setigerous, with second pair of branchiae also on posterior margin, dorsal to notosetae ( Fig. 1 View FIGURE 1 A–B); by about setiger 20, branchiae shifting to a more medial location and continuing to near posterior end ( Fig. 1 View FIGURE 1 A).
Parapodia of anterior and middle segments reduced to low ridges or mounds from which setae arise; posterior setigers modified with swollen podia bearing raised membrane from which acicular spines arise forming prominent cinctures. Anterior setae all heavy broad capillaries numbering 8–10 in noto- and neuropodia with notosetae longer than neurosetae; capillaries of middle segments, 40–48 in neuropodia and 49–55 in notopodia, similar in number but 1–2 becoming thicker, narrowing abruptly to thin pointed capillary tip; transitioning in posterior segments to blunt-tipped acicular spines; long, thin, natatory-like capillaries absent, including in mature specimens. Neuropodial blunt-tipped acicular spines first appear in posterior body segments from setigers 74 in the holotype and 68–84 in paratypes; notopodial spines from setigers 82–98. Noto- and neuropodial spines initially numbering 2–3 in a fascicle with 5–6 capillaries, then increasing to 3–4 spines; posterior full cinctures from last 6–8 setigers with up to 7–8 spines in noto- and neuropodia, with 15–16 spines on a side ( Fig. 2 View FIGURE 2 C), each spine alternating with a thin capillary seta ( Fig. 2 View FIGURE 2 D). High membranes of cinctured segments extending over dorsal midline forming shallow channel ( Fig. 2 View FIGURE 2 C); ventral surface flattened. Individual spines weakly curved, with narrow bluntly pointed tip; each with narrow internal core clearly visible in light microscopy ( Figs. 1 View FIGURE 1 D, 2D); spines appearing smooth externally in SEM ( Fig. 2 View FIGURE 2 E). Body terminating in simple pygidium bearing short triangular lobe ventral to anal opening ( Figs. 1 View FIGURE 1 C, 2F).
Methyl Green staining pattern. Numerous subdermal glands are concentrated on the prostomium, peristomium and in groups or bands along most of the body. These glands stain intensely with Methyl Green, creating a spectacular and distinctive staining pattern for this species ( Fig. 3 View FIGURE 3 B–E). Except for the anterior tip, the prostomium stains intensely on all sides ( Fig. 3 View FIGURE 3 B, E); the peristomium is intensely stained on the dorsal and lateral sides, with the dorsal stain extending along dorsal crest onto the anterior margin of segment 1 ( Fig. 3 View FIGURE 3 B, E). The less intense stain on Fig. 3 View FIGURE 3 B compared with Fig. 3 View FIGURE 3 C is due to a longer period of differentiation and destaining to show areas where the stain is most intense and retained for the longest period of time. Along the body, the segments anterior to the swollen area have dorsal and lateral bands of stain; the lateral bands are located in the middle of each segment, anterior to the setal fascicle ( Fig. 3 View FIGURE 3 E); dorsal to the notosetae, the stain expands into a prominent patch ( Fig. 3 View FIGURE 3 B–C) which gives way to transverse intersegmental bands that cross the dorsum ( Fig. 3 View FIGURE 3 B–C); between each of the dorsal bands are groups of stained glands producing a speckled pattern; ventrally, the banding is less prominent ( Fig. 3 View FIGURE 3 E). The swollen area includes all of the features of the preceding segments except that entire area is more darkly stained, producing the most intense and obviously stained part of the body ( Fig. 3 View FIGURE 3 B–D). Posterior to the swollen area, the staining pattern is similar to that of the anterior segmental area, but less intense. In the abdominal region, the stain is concentrated in thin transverse intersegmental bands and lateral longitudinal lines ( Fig. 3 View FIGURE 3 D).
Biology and ecology. Several paratypes are females with oocytes packed together in various stages of maturation; the largest eggs observed were 134–140 µm in the widest dimension. Specimens live in a muddy habitat (median grain size = 25 µm; 1.08% of organic carbon) offshore of the Gironde estuary ( 60 km) ( Masse et al. 2016). This species is important (13% of total abundance) in the local benthic community, which is dominated by polychaetes (16 species belonging to 14 families).
Remarks. Chaetozone elakata n. sp., with its fusiform shape due to expanded anterior body segments, is most similar to C. gibber Woodham & Chambers, 1994 from the NE Atlantic and Mediterranean Sea, C. hartmanae Blake, 1996 from off California in shelf and slope depths, C. larae from the SE Atlantic off Argentina in shelf depths, and to some extent C. bathyala Blake, 2015 from eastern Canada.
Chaetozone gibber differs from C. elakata n. sp. in having the expanded segments only dorsally enlarged providing a humped shape to the body instead of a more complete fusiform shape. In addition, the posterior cinctures of C. gibber are only partially developed with eight spines on a side instead of up to 16. The first segment of C. gibber is the first setigerous, whereas in C. elakata n. sp. the first segment is an achaetous segment anterior to setiger 1. Methyl Green stain has not been tested on C. gibber .
Chaetozone hartmanae has a similar fusiform shape as C. elakata n. sp., but the posterior cinctures have fewer than 10 spines on a side and, more importantly, the noto- and neuropodial spines differ in morphology with the notopodial spines being straight rather than curved and the neuropodial spines having a curved apex with serrations on the convex side ( Blake 1996). Additionally, the first segment of C. hartmanae is the first setigerous and also bears the first branchiae. The Methyl Green staining pattern in C. hartmanae is reduced to a band of speckles on the anterior and posterior margins of each segment.
Chaetozone larae View in CoL is similar to C. elakata View in CoL n. sp. in having a similarly expanded fusiform shape to the anterior body segments, a peristomium divided into two annular rings, one long and one narrow, and with both surmounted by a low dorsal crest. Chaetozone larae View in CoL and C. elakata View in CoL n. sp. also have a separate achaetous segment following the peristomium from which the dorsal tentacles arise. However, C. larae View in CoL is reported to also have the first pair of branchiae on the achaetous segment instead of setiger 1 as in C. elakata View in CoL n. sp. In addition, C. larae View in CoL has posterior cinctures with 11–13 spines on a side whereas C. elakata View in CoL n. sp. has posterior cinctures with 15–16 spines on side. In addition, the neuropodial spines generally begin more anteriorly in C. larae View in CoL , from setigers 43–74 versus setigers 68–84 in C. elakata View in CoL n. sp.; however, the segment where the spines begin is typically size related. The illustrations of the spines of C. larae in Elias et al. (2016) View in CoL suggest that they are shorter and with more of a curvature than those of C. elakata View in CoL n. sp. The most distinctive difference between these species, however, is with the Methyl Green staining pattern: C. larae View in CoL has no defined pattern, whereas C. elakata View in CoL n. sp. has numerous subdermal glands that stain with Methyl Green and produce distinctive staining patterns along the entire body. Chaetozone bathyala View in CoL has weakly expanded anterior segments, but this area is not prominent and does not resemble the swollen segments of C. elakata View in CoL n. sp. The presence of a complete post-peristomial achaetous segment preceding the first setigerous segment has been reported for several North American species:
Chaetozone bathyala View in CoL , C. careyi Blake, 2015 View in CoL , C. malmgreni Blake, 2015 View in CoL , C. palaea Blake, 2006 View in CoL , C. pigmentata Blake, 2015 View in CoL , and C. pugettensis Blake, 2015 View in CoL . Blake (2015) provided comparative details of these species; none of these species has a large swollen anterior region similar to that of C. elakata View in CoL n. sp.
Etymology. The epithet is derived from elakate, Greek for spindle, referring to the distinctive spindle or fusiform shape of the anterior segments of these worms.
Distribution. Bay of Biscay, 60 m in muddy sediments.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chaetozone elakata
| Lavesque, Nicolas 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
Chaetozone elakata
| Lavesque 2017 |
Chaetozone elakata
| Lavesque 2017 |
Chaetozone elakata
| Lavesque 2017 |
C. elakata
| Lavesque 2017 |
Chaetozone larae
| Elias, Rivero & Orensanz 2016 |
Chaetozone larae
| Elias, Rivero & Orensanz 2016 |
C. larae
| Elias, Rivero & Orensanz 2016 |
C. larae
| Elias, Rivero & Orensanz 2016 |
C. larae
| Elias, Rivero & Orensanz 2016 |
C. larae in Elias et al . (2016)
| in Elias et al. 2016 |
Chaetozone bathyala
| Blake 2015 |
Chaetozone bathyala
| Blake 2015 |
C. careyi
| Blake 2015 |
C. malmgreni
| Blake 2015 |
C. pigmentata Blake, 2015
| Blake, Achaetous Segment 2015 |
C. pugettensis
| Blake 2015 |
C . palaea
| Blake 2006 |