
Cryptocellus icamiabas, Tourinho, Ana Lúcia & Azevedo, Clarissa Salette De, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.177832 |
|
DOI |
https://doi.org/10.5281/zenodo.6249489 |
|
persistent identifier |
https://treatment.plazi.org/id/03D08787-FFD0-3A73-FF73-4EF144142353 |
|
treatment provided by |
Plazi |
|
scientific name |
Cryptocellus icamiabas |
| status |
sp. nov. |
Cryptocellus icamiabas View in CoL sp. nov
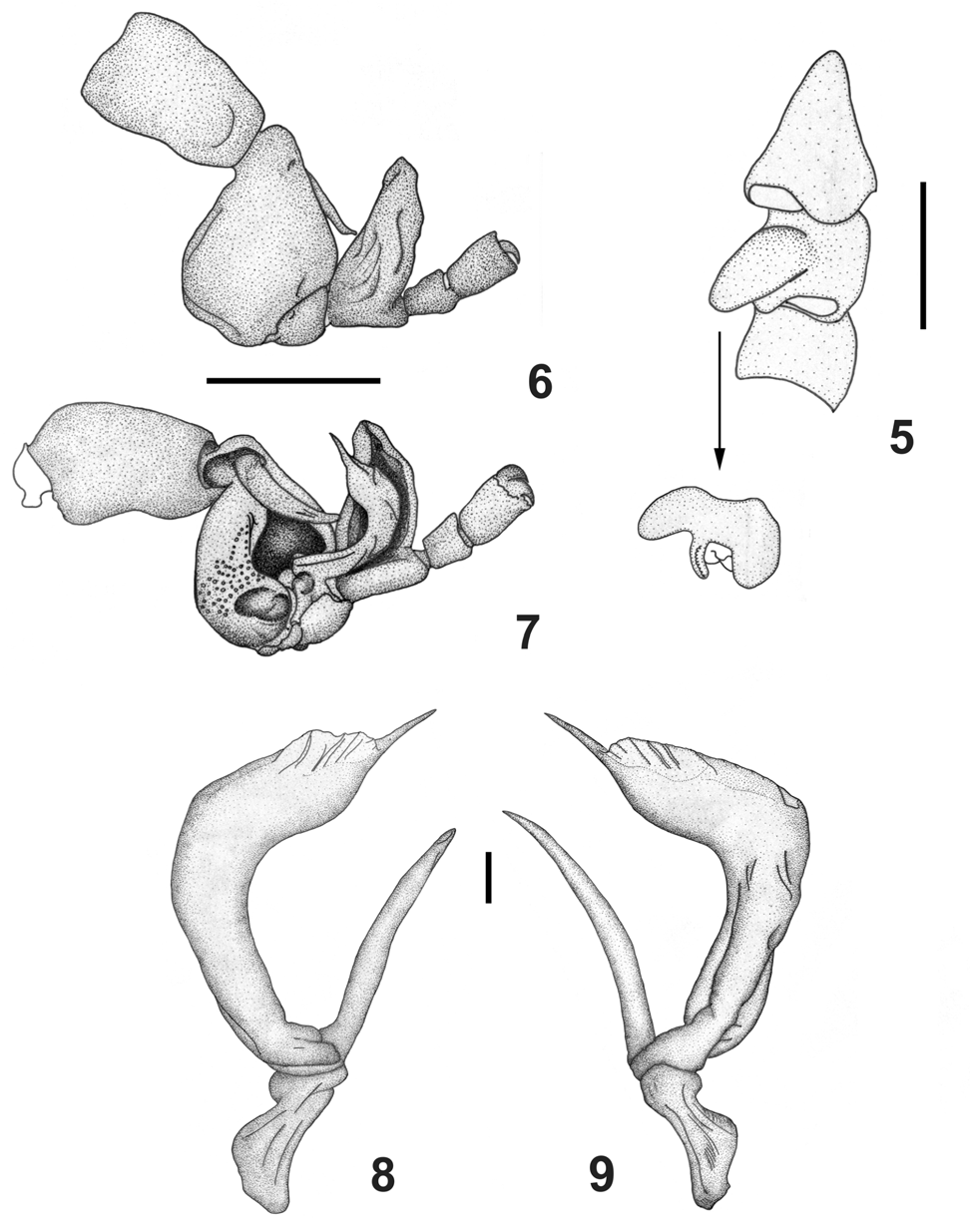
( Figs 1–10 View FIGURES 1 – 4 View FIGURES 5 – 9 View FIGURE 10 )
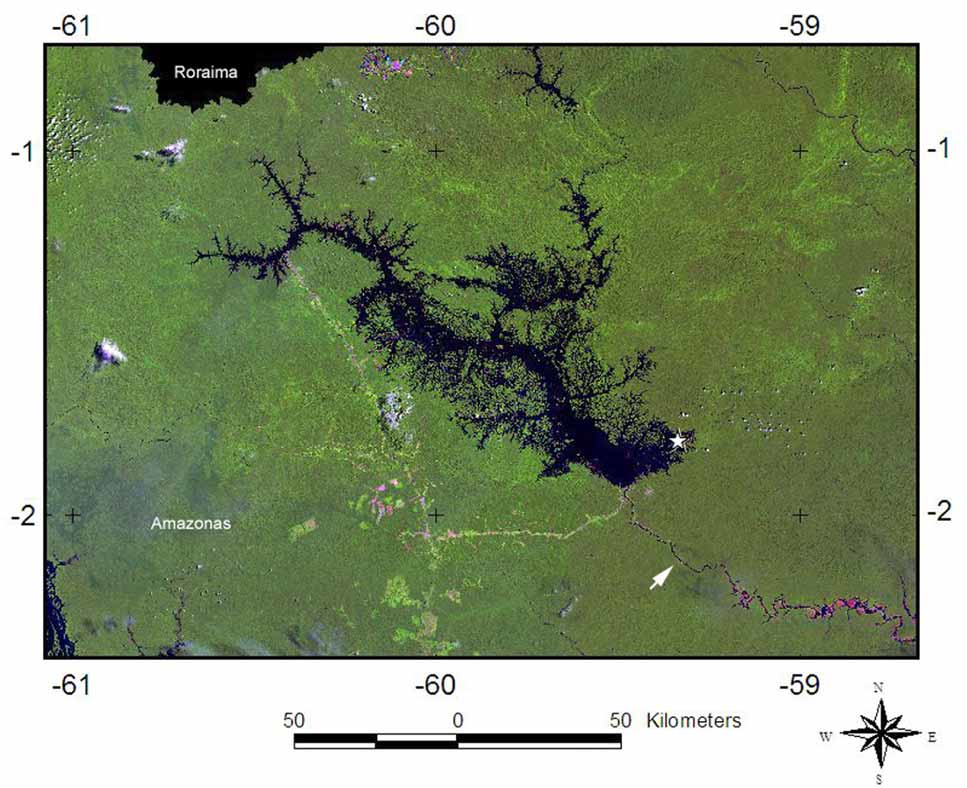
Type material. Male holotype ( INPA 10), Brazil, Amazonas State, Presidente Figueiredo, Balbina, 59,33117ºS - 1,79239ºW, 23.xii.2006, visual nocturnal search, transect 2, Ana Lúcia Tourinho leg.
Etymology. A noun in apposition that refers to the “ Icamiabas ”, the female warriors of Amazonian mythology described in the chronicles of the Dominican friar Gaspar de Carvajal. They have been identified as the Amazon warriors from Capadocia, from which the name of the Amazonas River and the Brazilian state of Amazonas derive.
Diagnosis. Cucullus with some much longer white setae on posterior margin only (fig. 2); ventral opisthosoma with a pair of median pits containing tubercles, there with darker and smoother tegument (fig. 3); tergal tubercles fewer than in C. abaporu , C. foedus Westwood, 1874 or C. simonis (fig. 4, see Platnick & Shadab 1977: figs 10, 22; Bonaldo & Pinto-da-Rocha 2003: fig. 9), but more numerous than in C. becki (see Platnick & Shadab 1977: fig. 44); margins of tergites slightly depressed and not tuberculate, tubercles mostly concentrated at the middle region (fig. 4). Proximal telotarsus II much more concave than in C. abaporu , C. becki or C. simonis (fig. 6); trochanter IV with a larger and rounded retrolateral apophysis (fig. 5).
Description of male holotype. Body total length, excluding pygidium, 3.9, cucullus 0.5 long, greatest width 1.1, prosoma 2.9 long, 2.5 wide between legs II and III; opisthosoma 3.0 long; 2.5 wide near middle of tergite; legs I 5.7, II 9.15, III 6.9, IV 6.8. General body color (in 70% ethanol) dark red, opisthosoma lighter than carapace, intersegmental membranes orange; Cucullus dark red, much darker at anterior margin and median half. Opisthosoma, legs and cucullus covered with rounded tubercles; iridescent tubercles, showing shades of purple and dark green, present on: prosoma (fig. 1), opisthosoma (more numerous), ventral opisthosoma (fig. 3), and legs I-III-IV. Surface of body entirely covered by setae concolourous with the body; prosoma with both straight and curved setae on anterior half, setae longer anteriorly and at posterolateral margins (fig. 1). Prosoma longer than wide, with some median tubercles, long and short setae on posterolateral sides (fig. 1). One pair of lateral eyes (fig. 1). Cucullus wider than long; concave between strongly protuberant lateral lobes, only tuberculate in this concavity, proximal margin convex medially; with large and deep anterior depression, densely covered with long white setae (fig. 2). Chelicerae as in C. abaporu . Sternal region with coxa I not meeting tritosternum; left and right coxae II meeting in midline anteriorly and posteriorly, but not in-between; coxae III meeting; coxa IV meeting anteriorly; midline suture of coxae III slightly longer than those of coxae II and IV. Opisthosomal tubercles spread on lateral tergites, intersegmental membranes without tubercles or setae ventrally: only present on posterior half of opisthosoma, mostly concentrated at lateral concavity and posterior margin; no median tubercles. Pygidium with very distinct distal dorsal notch on basal segment; no ventral notch. Pedipalps orange, without tubercles. Distal leg coxae darker, leg tarsi light red. Leg formula II>IV>III>I. Small and round prolateroventral apophysis in trochanter of leg III; teleotarsus II of tarsus III, in anterior view, with two connected inner apophyses (fig. 7). Copulatory apparatus as illustrated (fig. 8–9). Legs with numerous sexual modifications. Femur III with numerous tubercles; apophyses of trocanter III as in C. abaporu . Leg I as in C. abaporu , metatarsus I with small ventral protuberance; proximal end of femur II with slight protuberance only; trochanter and femur III as in C. abaporu .
Female. Unknown.
Distribution. Known only from the type locality (fig. 10).
Remarks. Some structures of C. icamiabas give the impression of a morphological mosaic of two, three and sometimes even four different species. C. icamiabas seems to be closest to C. abaporu , although in C. icamiabas the copulatory apparatus (figs 7–9), regarded as the most useful taxonomic character ( Selden 1992), is similar to those of C. abaporu and C. simonis . The apophyses of the trochanter of leg III are similar to those of C. abaporu and C. whitticki , with the prolateroventral apophyses being smaller and more rounded than in C. simonis and C. becki . The apophysis on the trochanter of leg IV is as long as that of the first two species, but is rounded (fig. 5), rather than pointed. Tarsus III very much as in C. abaporu , C. becki and C. simonis , except for the wider telotarsus II; in anterior view, the two connected inner apophyses seem to be represented by just one in C. simonis , (fig. 6–7, see Platnick & Shadab 1977: fig. 24), the basitarsus, in posterior view, does not show the convex shape seen in C. abaporu . The distal copulatory apparatus of C. icamiabas (figs 7–9) is closer in form to that of C. simonis , while the proximal end is very much as in C. abaporu . In general, the copulatory apparatus is quite similar to that of C. abaporu , C. whitticki and C. becki . The density of the tergal tubercles is lower than that seen in C. abaporu , C. foedus and C. simonis (several), but higher than that observed in C. becki (very few). The interrelationships between C. icamiabas and the other species will only become clear when the female of the former is found.
Notes on the Biotope. Balbina Lake is the reservoir of the Balbina hydroelectric power plant, which formed as a result of the construction of a dam across the Uatumã River in 1987. This gave rise to island assemblages bearing remnants of the original forest. Most of the vegetation on these islands was burned about ten years ago by an accidental fire caused by the illegal activities of local fishermen, which is one of the reasons why the vegetation is now mainly composed of lianas, and young palms and other trees. Some of Lake Balbina’s islands are permanently isolated, while others are only periodically separated by the seasonal pulse of water in the Amazon. These different factors have a large influence on the composition, abundance and richness of the species living on the islands.
Despite the negative environmental impact of the dam’s construction, the islands of Lake Balbina and the continous forest area of the Uatumã Reserve are important areas for the study of arachnids, showing a high species richness and an impressive number of undescribed taxa of Araneae and Opiliones, as well as providing new records of genera and families for the Amazon basin ( Ferreira 2007). The holotype of C. icamiabas was collected at night by visual searching, when it was found resting on the soil surface. Because the project was originally designed for a rapid analysis of the richness and composition of the spider fauna, no specific methods for catching ricinuleids (e.g. Winkler or Kempson extraction) were applied. Leaf litter was manually inspected during night searching (30 meters transects, one hour searching), and manual litter sorting was done during the day (three samples of litter were collected, each 10 meters apart, along a transect of 30 meters).

| INPA |
Instituto Nacional de Pesquisas da Amazonia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |