
Trogontherium (Euroxenomys) minutum rhenanum Franzen, Storch, 1975
|
publication ID |
https://doi.org/ 10.1515/vzoo-2015-0062 |
|
persistent identifier |
https://treatment.plazi.org/id/03CEDC4B-EB66-DE5A-FF05-F968FC869691 |
|
treatment provided by |
Felipe |
|
scientific name |
Trogontherium (Euroxenomys) minutum rhenanum Franzen, Storch, 1975 |
| status |
|
Trogontherium (Euroxenomys) minutum rhenanum Franzen, Storch, 1975
Monosaulax sp. : Topachevsky et al., 1987, 1992, 1997; Steneofiber sp. : Nesin, Nadachowski, 2001; Euroxenomys minutus rhenanus (comb. n.): Casanovas-Vilar, Alba, 2011.
T y p e l o c a l i t y. Dorn-Dürkheim, Germany.
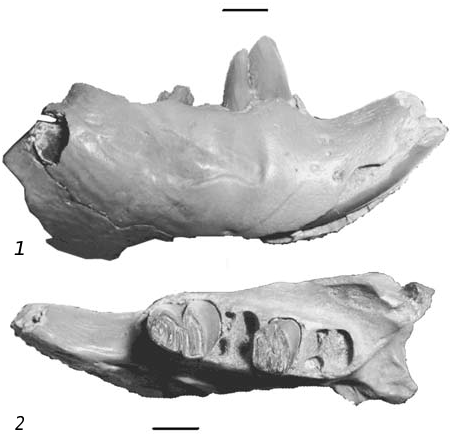
I n v e s t i g a t e d m a t e r i a l. Mikhailovka 2 (MN 11): 1 M 1-2, dP 4; Popovo 3 (MN 12): 1 dP 4; Cherevichnoe 3 (MN 12): 1 P 4, 1 dP 4, 1 mandible with P 4 -M 2 ( fig. 2 View Fig ), 2 M 1-2, 1 P 4 ( fig. 3 View Fig , 8), 5 M 1-2, 3 M 3; Belka (MN 12): 1 M 3; Nikomarovka: 1 M 1-2; Novoelizavetovka 2: 1 M 1-2; Vinogradovka 1: М 1-2 — 5, М 3 — 2, Р 4 — 3, М 1-2 — 7; Pontian lectostratotype: Р 4 — 5, М 1-2 — 2, М 3 — 3, Р 4 — 3, М 1-2 — 5; Andreevka: Р 4 — 1, М 1-2 — 2, М 3 — 2, Р 4 — 2, М 1-2 — 4.
G e o l o g i c a l a g e: MN 10–13, Late Miocene (Turolian).
The earliest representatives of the T. (Euroxenomys) minutum rhenanum in Ukraine are known from the Mikhailovka 1 and 2 locality.
D e s c r i p t i o n a n d c o m p a r i s o n. P 4 from the numerous localities on the territory of Ukraine ( fig. 3 View Fig , 1 View Fig ) are characterized by a somewhat larger size as compared to the nominative subspecies and those from Dorn-Dürkheim. The height of the tooth crown is on average 4.2 mm (maximum — up to 8.0 mm), length — within the range of 3.6–5.3 mm, width — 4.1–6.1 mm, and W/L ratio is 110 % (table 1). The widest teeth are observed in materials from Mikhailovka on Bug 2 and Verkhnya Krynitsa 2 (W/L ratio — 115.0 %). Hypostria usually reach the crown bottom, mesostria is long; flexus closed into the fossettes in early stages of wear; parafossette and metafossette are insular-shape. Tooth size of the subspecies from Dorn-Dürkheim is similar, but the latter have almost the same length and width (W/L ratio — 98 %). Length of the dP 4 from Vinogradovka 1 is 2.9 mm, width — 3.0 mm (W/L = 103 %), the height of the tooth crown is 4.6 mm. Paraflexus tends to branching, mesoflexus crosses the entire width of the chewing surface, metafossette is circular. Apex of the hypoflexus reaches the middle part of the chewing surface, para- and mesostriae are weak, hypostria reaches the crown bottom.
Height of M 1-2 from Andreevka, Vinogradovka 1 and Pontian lectostratotype is on average 8.0 mm, length — near 3.5 mm, width is 4.0 mm (W/L ratio — 114 %). Parafossette and mesofossette are well-expressed ( fig. 3 View Fig , 2 View Fig , 3 View Fig ); hypofossette is shifted to the labial edge, while its lingual side docks with the labial edge. Parafossette is sometimes small and short in buccal-lingual direction, hypostria ends below half of the crown height. Insular-shape hypofossette is often small, slightly elongated, and located parallel with the parafossette. Paraflexus reaches half of the chewing surface width in contact with hypoflexus. Lingual edge of mesoflexus almost reaches the edge of the chewing surface. Metafossette is oval. Meso- and metastries are weak and hardly noticeable. One M 1-2 from Frunzovka 2 is slightly different in size and morphology: L = 5.5 mm, W = 3.8 mm (W/L ratio — 69.1 %), i. e. the tooth is longer than wider.
Elongated M 3 with well-developed roots from Belka, Andreevka ( fig. 3 View Fig , 4), Pontian lectostratotype are relatively large (L in the range of 4.0– 5.5 mm, W = 3.3–4.0 mm, the average index — 75 %). Parafossette is located transversely with hypostriid, mesostria is long, ends by less than half of the crown height. Metafossette is Y-shaped; there are two small additional fossetids. There is a small additional fossette under the parafossette, from the labial tooth side; mesoflexus is not closed, there are two small fossettes at the bottom of metaflexus. Hypostria reaches extends about nearly 2/3 of the crown height. Teeth from the Pontian lectostratotype have an oblong parafossette and a long arched mesofossette. Metafossette disposed substantially parallel to the longitudinal axis of the chewing surface. There is an additional (fourth) fossette inside the bend of metafossette.
M 3 from Belka is shorter and wider than those from Dorn-Dürkheim, while inferior in the size of the teeth from the Pontian lectostratotype. M 3 from Andreevka is smaller than those from the Pontian lectostratotype and Dorn-Dürkheim. M 3 length from Vinogradovka 1 is 3.0 mm, width — 2.2 mm, crown height is 6.4 mm. The tooth is slightly worn; paracone is insular (isolated). Parafossette in the teeth from the Pontian lectostratotype is located on the labial tooth side; mesofossette is expressed across the entire width of the chewing surface and permanently bonded to the front part of the tooth. The metafossette is slightly curved. Additional fossettes, different in form, are located down. Hypostria reaches 1/3 of the crown height.
P 4 morphology of the studied subspecies from the Late Miocene localities are divided into two groups — early or small, including the remains from MN 11–12, and late or larger — MN 13. Differences between them are stored mainly in terms of size. Tooth length of the early Maeotian (MN 11–12) populations is in the range of 5.1–5.7 mm, width — 4.1–4.6 mm, W/L ratio is 80 %. The larger tooth size is characteristic for the populations, dated by the Pontian (MN 13): 6.0 × 4.3 mm in average, W/L ratio — 71.5 %. Beavers from Andreevka, Vinogradovka 1 and Pontian lectostratotype have less broad teeth and reflect a general trend to P 4 elongation in time. The population from Dorn-Dürkheim takes an intermediate position: under size (6.2 × 4.8 mm) it is closer to those from the Pontian lectostratotype; however under the W/L ratio (77 %) it is similar to Maeotian populations. Height of the tooth crown is in the range of 8.0–9.0 mm, roots are moderately developed. Parafossetid is elongated and slightly curved, mesoflexid is straight, and metafossetid is located on the labial tooth side and similar in shape and size to parafossetid. Hypoflexid abuts by the labial edge into the lingual edge of metafossetid. Mesostriid reaches 2/3 of the crown height. Occasionally there is an additional rounded fossetid between para- and mesoflexids. A slightly different P 4 morphology is peculiar for the remains from Andreevka ( fig. 3 View Fig , 5). Tooth length is 6.7 mm, width — 4.5 mm (W/L index — 67 %). There is an additional fossetid in the front part of the tooth; other fossetids have approximately the same shape. Hypoflexus comes on the chewing surface between the mesofossetid and metafossetid; hypostriid reaches the bottom of the crown, and the height of the crown is 5.2 mm. Tooth roots are greatly developed, exceeding the crown height.
M 1-2 of this subspecies from the Maeotian localities of Ukraine belongs to adult individuals ( fig. 3 View Fig , 6, 7). They have poorly developed roots; there is no cement at the crown base. Crowns are slightly curved, their average height is 8.0 mm, hypostriid length — 3.9 mm, parastriid height is 0.1 mm, mesostriid — 1.2 mm. Metafossetid is somewhat curved and well-expressed. There is a fourth (additional) fossetid that distinguishes it from other Palearctic subspecies. The tooth crown is almost square (W/L ratio is in the range of 97–100 %). Parafossetid on individual teeth, e. g. from the Verkhnya Krynitsa 2, shifted leftwards, mesoflexid comes under the lingual edge of the hypoflexid, metafossetid is well-expressed. Hypostriid lowered on ¾ of the crown height. Mesoflexid and metaflexid formed two (lingual and labial) striids in the process of crown wear. There is a small rounded fossetid below the metaflexid, in the middle part of the lower edge of hypoconid. Labial edge of the mesostriid is weakly expressed, lingual edge is very small. Metastriids and metaflexies are also hardly expressed. Hypostriid reaches more than half of the crown height.
Remains of T. minutum minutum from Grytsev (MN 9) are similar to those from the Höwenegg locality in Germany ( Giersch et al., 2010) in size of the lower molars as well as tooth proportions. Subspecies of Grytsev locality also has similar size with Euroxenomys minutus minutus from Sansan, but differs by narrower teeth, especially in the lower jaw, and relatively broader and shorter P 4 (109 % against 96.0 % for the subspecies from Sansan). Remains of T. minutum ozansoyi are closest to subspecies from Sansan in size and proportions of P 4 and M 1-2 (table 1). This group of remains includes the most ancient and most minor forms of the nominative subspecies; they differ from the later and larger subspecies T. minutum rhenanum .
All teeth from the Maeotian localities are somewhat smaller as compared to those from Dorn-Dürkheim , Andreevka , Vinogradovka 1 and Pontian lectostratotype (table 1). Besides , typical series has wider teeth (W/L index — 122.0 %) unlike the Maeotian forms. M 1-2 are smaller in Maeotian localities (average length is 3.4 mm, width — 3.35 mm, W/L ratio is 97.0 %), i. e. teeth are clearly square. Teeth from the late Maeotian and Pontian localities (Andreevka, Vinogradovka 1, Pontian lectostratotype) are larger (average length is 3.5 mm, width — 4.4 mm, W/L ratio is 140 %), i. e. width is much greater than length. It is an important indicator, especially for the remains from the Vinogradovka 1. Height of the crowns is in the range of 6.0–8.0 mm. T. (Euroxenomys) minutum rhenanum from Ukraine differs under the marked morphological features from the type series of Dorn-Dürkheim, which is closer to the Pontian populations. M 1-2 from Andreevka are longer, but narrower than those from the Pontian lectostratotype. Besides, they are larger than those from Novoukrainka, Nikomarovka, and Verkhnya Krynitsa 2. M 1-2 from Andreevka is bigger than those from Dorn-Dürkheim, and wider than longer .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.