
Bothriolepis prima Gross, 1942
|
publication ID |
https://doi.org/ 10.5281/zenodo.4664755 |
|
DOI |
https://doi.org/10.5281/zenodo.10248681 |
|
persistent identifier |
https://treatment.plazi.org/id/03CC6624-FFA9-FFE2-FC9E-FE1D9DADFA8F |
|
treatment provided by |
Felipe |
|
scientific name |
Bothriolepis prima Gross, 1942 |
| status |
|
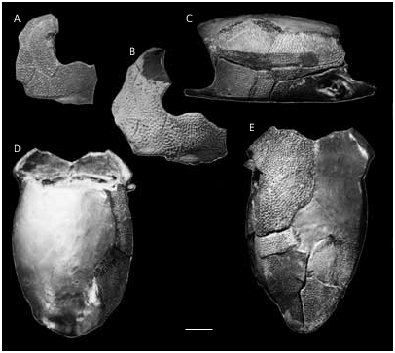
Bothriolepis prima Gross, 1942 ( Figs 4-6 View FIG View FIG View FIG )
Bothriolepis prima Gross, 1942: 415 , 416, abb. 6A.
HOLOTYPE. — AMD, Riksmuseet, Stockholm.
MATERIAL EXAMINED. — Disarticulated plates ( LGI 5/1064, 1066, 1072-1074, 1080, 1089, 1215, 2202-2204, 2243, 2245-2247, 2253, 2254, 2264-2270, 2272, 2273, 2286, 2295-2299, 2301, 2302, 2304, 2305, 2307-2311, 2322, 2323, 2325-2327, 2341, 2342, 2383, 2385, 2388, 2398-2406, 2410-2412, 2699-2703). Fragments of the trunk-shield and pectoral fin bones ( LDM 43/584) are specimens additional to the material described by Karatajūte-Talimaa (1966) from Pastamuiža locality.
LOCALITIES AND HORIZON. — Daugava River near Pastamuiža in the vicinity of Koknese (Kokenhusen), Latvia; Upper Devonian, lower Frasnian, upper part of the Amata Formation. Pelyša River, Lithuania; uppermost part of the Šventoji Formation. YamTesovo at Oredezh River, Leningrad region of Russia; Staritsa Beds of the Yam-Tesovo Formation.
DIAGNOSIS. — Small Bothriolepis with a median dorsal armour length reaching 50 mm. B/L index of the head-shield of 140-150. Weakly convex rostral margin slightly shorter than the posterior margin; obtected nuchal area present only on Nu. Orbital fenestra is relatively large. Nu bears posterior process. Dorsal wall of trunk-armour broad and high in its anterior part and relatively narrow in posterior part, B/L index 77. Lateral wall is high. Tergal angle and median dorsal ridge are weakly defined. Postlevator cristae on the visceral surface of AMD are straight and enclosing a relatively sharp angle. AMD relatively broad, arched, B/L index of 99. Dorsal lamina of ADL relatively narrow, dorsal and lateral laminae enclosing an angle about 122-128°. Angle between the dorsal and lateral walls on MxL is sharper: about 112°. Lateral and ventral walls enclosing an angle about 109° on PVL. Dorso-lateral ridge is weakly defined in its anterior part and is very well-developed in the posterior part of the trunk-shield. Ventro-lateral ridges are strongly developed. Lateral lamina of PVL is 1.7 time as long as it is high. Ventral wall of the trunk-shield is relatively narrow, B/L index of 46, with relatively broad anterior part and narrow posterior part. Lateral and mesial spines on the proximal segment are separate and relatively long. Ornamentation reticulate in small and moderately large individuals, tuberculate in large individuals. Central part of the dorsal and ventral walls is almost smooth. Pit-line groove crossing the dorso-lateral ridge on the MxL is always present.
DESCRIPTION
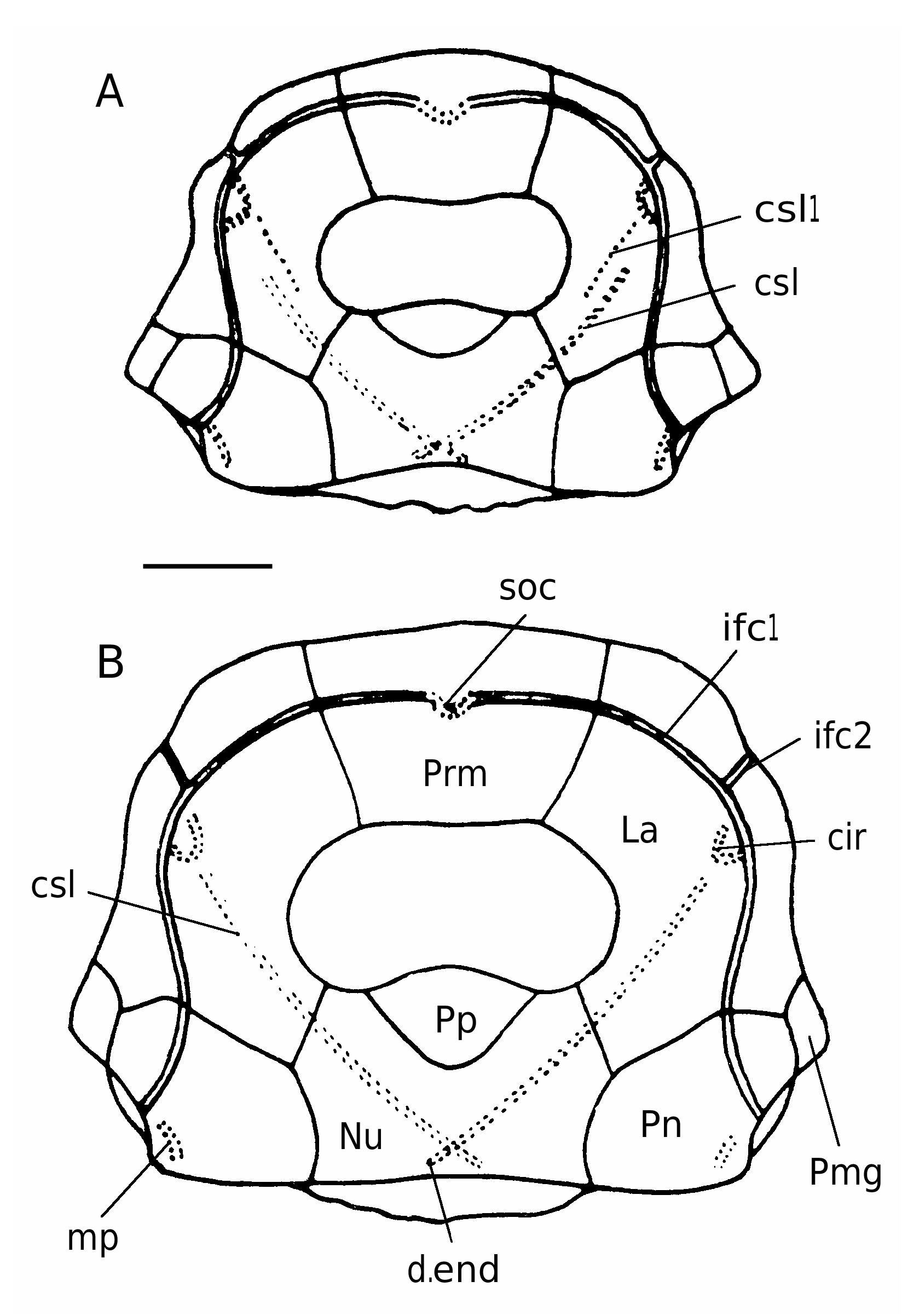
As noted by Karatajūte-Talimaa (1966), the head-shield ( Figs 4A, B View FIG ; 5 View FIG ) is strongly vaulted and broad. The antero-lateral corners (alc) and the prelateral notch (nprl) are gently defined. The obtected nuchal area (nm) is relatively broad, well-defined only on the Nu. The orbital fenestra is large, but relatively narrow. Such proportions of orbital fenestra in antiarch fishes is typical for young individuals ( Long & Werdelin 1986; Upeniece & Upenieks 1992).
The Prm is moderately broad and short, it is broadest at the antero-lateral corners of the plate. The rostral margin is slightly convex. The orbital margin is concave in specimen LGI 5/1064 from Pelyša River, it is only slightly shorter than the rostral margin ( KaratajūteTalimaa 1966). The infraorbital sensory groove crosses the plate in its anterior part, but not so close to the rostral margin as in B. obrutschewi (pers. obs.).
The La, Pp and Nu were not described by Karatajūte-Talimaa (1966) in details. The La is moderately broad with the L/B index 140-144, 142 on the average (n = 4). It is narrow in the anterior part and broad in the posterior part. The rostral margin is relatively short and almost straight, the antero-median margin is short. The infraorbital sensory line groove crosses the plate in its anterior part not far from the rostral and lateral margins. The central sensory line groove (csl) finishes at the level of middle of the orbital margin of the orbital fenestra. The Pp is broad in small individuals and of moderate breadth in maturity. Its anterior margin is convex in both small and larger specimens.
The Nu is relatively broad with a L/B index 68- 70 (n = 3). The plate is usually broadest across the lateral corners, but in specimen LGI 5/2402 it is broadest across the postero-lateral corners. The anterior division of the lateral margin usually is straight and a little shorter than the strongly concave posterior division. The anterior margin with the deep and broad postpineal notch (Karatajūte-Talimaa 1966). The posterior margin is usually almost straight or slightly concave. The central sensory line groove is clearly distinct. The outer openings of the endolymphatic ducts are small but clearly defined, lying not far from the obtected nuchal area.
The Pn is of moderate breadth, L/B index is 74 and 83 in two measured specimens. The plate bears clearly seen middle pit-line groove (Karatajūte-Talimaa 1966).
The Pmg is broad with the lateral margin slightly longer than the median margin ( KaratajūteTalimaa 1966).
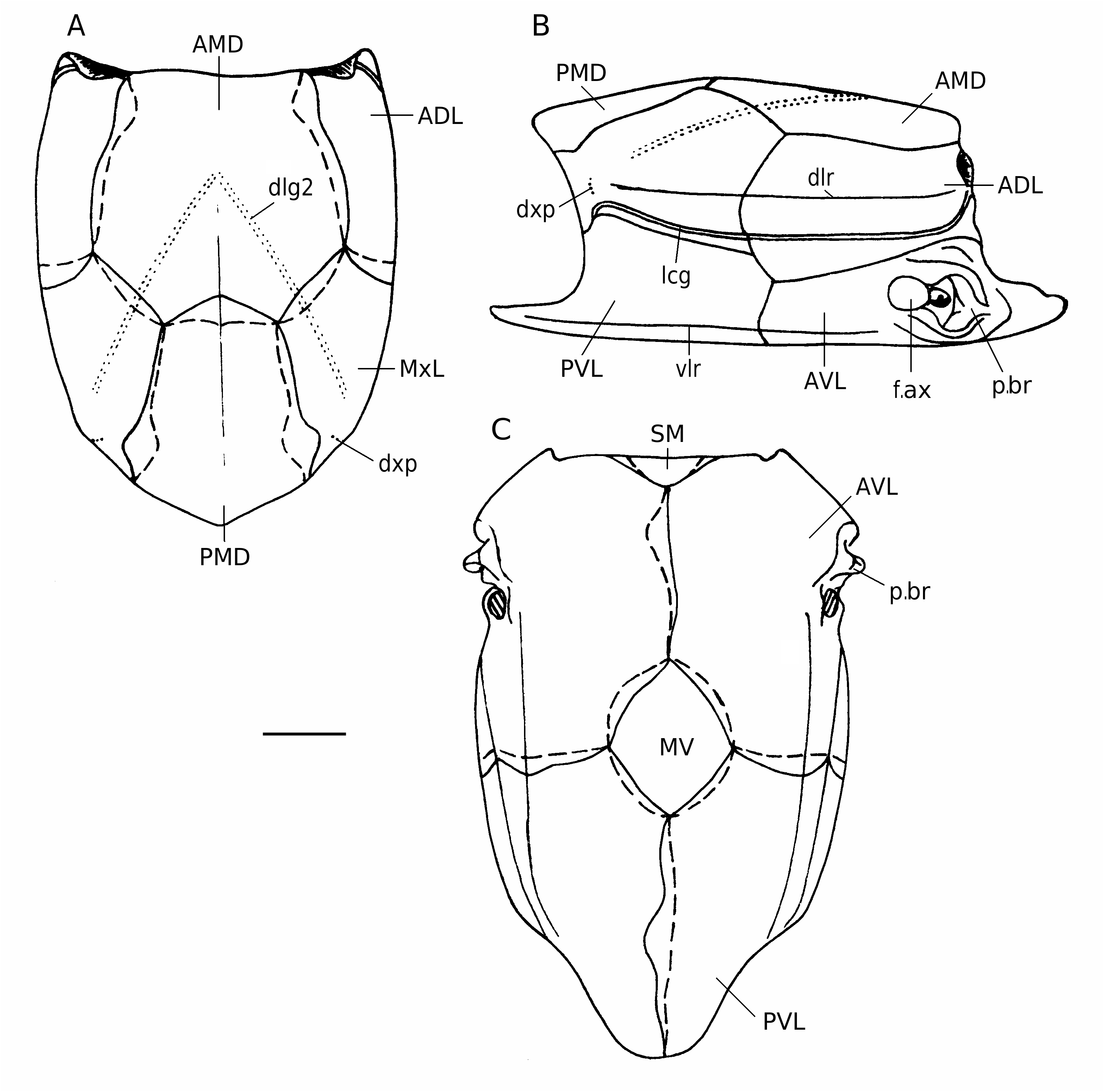
The trunk-armour ( Figs 4 View FIG C-E; 6) is relatively narrow as restored by Karatajūte-Talimaa (1966: figs 3, 4), with broader anterior part of the dorsal and ventral walls of the trunk-armour and much narrower posterior part of that walls, hence broad subcephalic division and narrow and relatively high apperture for the tail. Karatajūte-Talimaa suggested that this fish had narrower and shorter tail than the other representatives of the genus Bothriolepis . The length of the dorsal wall reaches about 60 mm. The dorso-lateral and ventro-lateral ridges are wellmarked, especially the ventro-lateral ridge and the posterior part of the dorso-lateral ridge. The median dorsal ridge is weakly defined, and is usually better developed posteriorly, on the PMD. The angle enclosed by the dorsal and lateral walls is obtuse in the anterior part of the trunk-armour (measured on the ADL: 122- 128°), and is more sharp in the posterior part (on the MxL: 112° on the average). The angle enclosed by the lateral and ventral walls is about 109° (all angle measurements follow KaratajūteTalimaa 1966).
The AMD is relatively broad, B/L index is 92 and 105 in two measured specimens, characterising by a variable shape. As noted by KaratajūteTalimaa (1966), the anterior margin is broad, weakly convex, posterior margin short, 1.2- 1.8 time shorter than the anterior margin. The antero-lateral and lateral corners are welldefined, but the postlevator processes are weakly developed. The posterior division of the lateral margin is only 1.2-1.9 time shorter than the anterior division (Karatajūte-Talimaa claimed that this feature is rather variable and reaches 1.13-3.22 in B. prima , but I did not find the specimens with such an extremely long anterior division of the plate among the material). The median dorsal ridge and the tergal angle (dma) are weakly marked. Overlap areas for the MxL are normally developed as usually in Bothriolepis ; only in one case for 48 overlooked by Karatajūte-Talimaa (1966), in LGI 5/2247, the AMD overlaps the MxL by its anterior part of the posterior division of the lateral margin similar as in Remigolepis . The anterior (dlg1) oblique dorsal sensory line grooves are weakly defined only on the plates of individuals of small size: LGI 5/2247 and 5/1066 with the length of the AMD 13 and 17 mm respectively (pers. obs.). The posterior (dlg2) oblique dorsal sensory line grooves usually are well-defined.
The visceral surface of the AMD shows a slightly lengthened triangular-shaped levator fossa (f.retr), which is limited by the low postlevator thickenings (alr) without the postlevator cristae (Karatajūte-Talimaa 1966), as it is seen in the specimens from Yam-Tesovo, or probably in a posterior part also by the postlevator cristae, as it was figured by Gross (1942: abb. 6A).
The PMD is relatively narrow, B/L index is 75 and 86 in two measured specimens. The posterior margin is usually strongly convex, but rounded, without pronounced posterior corner (Karatajūte-Talimaa 1966). The width of the anterior margin varies between 46-56% of total breadth, being almost of the same proportions as that of B. obrutschewi . The lateral corners are well-defined, the postero-lateral corners are often rounded. The median dorsal ridge is usually present only in the posterior half of the plate in well-grown individuals (pers. obs.).
The ADL is moderately broad, the dorsal lamina is 2-2.3 time as long as it is broad and its breadth 1.1-1.2 time exceeds height of the lateral lamina. The dorso-lateral ridge (dlr) is better defined in the posterior part of the ADL (Karatajūte-Talimaa 1966). Dorsal and lateral laminae of the plate enclosing an angle of about 124° on the average (Karatajūte-Talimaa 1966), which is more obtuse than that in B. obrutschewi . The processus obstans is strongly developed (Karatajūte-Talimaa 1966).
The MxL was not described by KaratajūteTalimaa (1966) in details except the proportions, angle between laminae, and sutural connections of the plate. The dorsal lamina of the plate is 1.4- 1.7 time as long as it is broad, 1.57 on the average, and possesses a well-defined relatively sharp dorsal corner. The lateral lamina is relatively low, the dorsal length of the plate 2.8-4.1 times (3.6 on the average) exceeds the height of the lateral lamina. The posterior oblique sensory line groove (dlg2) terminates close to the lateral margin, in some distance from the dorso-ventral pitline groove (dxp) crossing the dorso-lateral ridge, similarly to B. obrutschewi , but in specimen LGI 5/1072 both grooves are connected with each other. The overlap area for the AMD is usually as normally in Bothriolepis , only in one case ( LGI 5/2403) it is restricted to approximately one fourth of the length of the anterodorsal margin (Karatajūte-Talimaa 1966), as in Remigolepis ( Stensiö, 1931) .
The AVL is relatively broad in subcephalic division (Karatajūte-Talimaa 1966), the ventral lamina is 1.5-1.6 time (n = 3) as long as it is broad. The subcephalic division is extremely short and comprises only about 15% of total length of the ventral lamina. The antero-lateral corner is situated slightly medially the axis of the ventrolateral ridge. The ventral lamina is 2.6-3 times as broad as the low lateral lamina high. The axillary foramen (f.ax) is relatively large and rounded or gently elongated in shape (4.1 × 3.6 mm in LGI 5/2701 with total length of the ventral lamina reaching 33.3 mm).
The PVL is relatively broader than that in B. obrutschewi : the ventral lamina is 2.0- 2.3 times as long as it is broad. The subanal division occupies about one fourth or fifth (17-26%) of the total PVL length. The lateral lamina is high, the ventral lamina is 1.1-1.5 (1.28 on the average) time as broad as the lateral lamina high. The angle between laminae is more steep as in B. obrutschewi and reaches about 109° (Karatajūte-Talimaa 1966). The left PVL overlaps the right one by a wide overlapping area (Karatajūte-Talimaa 1966). The ventro-lateral ridge (vlr) is clearly defined, sometimes slightly pronounced over the lateral lamina.
The MV is typically developed: it is relatively small and of rhombic shape (Karatajūte-Talimaa 1966).
The pectoral fin is represented by several disarticulated bones, and three specimens showing articulated plates of the proximal segment. Karatajūte-Talimaa (1966) claimed that the L/B index of the proximal segment is 3.8-4.5, but I found it to be more elongated being 4-4.6 times as long as it is broad. The CV1 is making contact with the MM2. The distal segment is much shorter than that in B. obrutschewi , it is 3.8 times as long as it is broad ( KaratajūteTalimaa 1966). It shows no traces of an individual plates. Probably, Karatajūte-Talimaa (1966) correctly suggested that the pectoral fin does not extend posteriorly far from the posterior margin of the trunk-armour ventral wall. The CD1 is relatively short and broad with L/B index varying from 2.7 to 2.9 (2.8 on the average). The CV1 is slightly longer than the CD1. The ML2 is elongated, proportionally much longer than that in B. obrutschewi , it is 4.8- 5.1 times as long as it is broad measured from the dorsal side, and 4.1-4.7 times as long as it is broad measured from the ventral side. Both segments bear prominent lateral and mesial spines. The well-defined mesial edge of the proximal segment bears isolated mesial spines and the lateral spines are fused in their base forming welldefined crest (Karatajūte-Talimaa 1966).
The ornamentation was well described by Karatajūte-Talimaa (1966), it is typically reticulate, in small and medium-sized individuals of a fairly regular fine-meshed network of welldefined anastomosing ridges, becoming almost smooth in the middle part of the dorsal wall of the trunk-armour, including the posterior part of the AMD, the anterior part of the PMD, the postero-median part of the ADL, and the antero-median part of the MxL, as well as on the median part of the ventral wall. In rather large individuals ornamentation of the head-shield and dorsal wall of the trunk-armour become more tuberculate. The ornamentation on the subcephalic division of the AVL consists of oblique tubercular ridges, and sometimes there are several tubercular ridges parallel to the posterior margin on the subanal division of the PVL.
REMARKS
The following account is the first full treatment in English, which supplements KaratajūteTalimaa’s (1966) good description (in Russian) of Bothriolepis prima , adding some details of structure. It is based mainly on the collections of Karatajūte-Talimaa’s and Lyarskaja.
DISCUSSION
Bothriolepis prima resembles B. obrutschewi and B. evaldi from the Main Devonian field, and B. hydrophila ( Agassiz, 1844) ( Miles 1968) from Scotland. For differences with B. obrutschewi and B. evaldi , see the descriptions below. B. prima differs from B. hydrophila in 1) smaller size; 2) broader dorsal wall of the trunk-shield; 3) shape and proportions of the AMD; 4) narrower ventral wall of the trunk-shield; 5) shorter proximal segment of the pectoral fin. B. prima can be distinguished from B. cellulosa (see Gross 1941), other lower Frasnian Bothriolepis representative from the Main Devonian Field mostly by 1) its much smaller size; 2) narrower lateral division of the Pn; 3) proportions of the trunkarmour; 4) narrower PMD; 5) relatively narrow- er proximal segment of the pectoral fin; 6) CV1, which is making contact with the MM2; 7) smooth median part of the dorsal wall of the trunk armour. Small Bothriolepis remains of similar size from Kazakhstan ( Malinovskaya 1977, 1988) are too incompletely described and figured to make the detailed comparisons.
| LDM |
Latvian Natural Histotry Museum, department of Entomology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bothriolepis prima Gross, 1942
| Lukševičs, Erwin 2001 |
Bothriolepis prima
| Gross 1942: 415 |