
Caridina buehleri Roux, 1934
|
publication ID |
https://doi.org/10.5281/zenodo.176586 |
|
DOI |
https://doi.org/10.5281/zenodo.6249873 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB87FF-0013-FFFD-3B88-C223FC10FED9 |
|
treatment provided by |
Plazi |
|
scientific name |
Caridina buehleri Roux, 1934 |
| status |
|
Caridina buehleri Roux, 1934 View in CoL
( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 )
Material examined: One male cl 3.8 mm, rl 5.2 mm and 3 ovigerous females (eggs without eyes) cl 4.6–5.7 mm ( MZB Cru 1566), Indonesia, Central Sulawesi, Luwuk Peninsula, Freshwater spring in Malontong, west of Ampana, brackish water pool; 0°53.125’S 121°31.371’E (field code 70-05); coll. M. Glaubrecht, T. von Rintelen & K. Zitzler, May 27, 2005.1 male cl 2.9 mm, 1 female cl 3.9 mm and 3 ovigerous females (eggs without eyes) cl 4.3–6.4 mm ( ZMB 29002), same data as above.
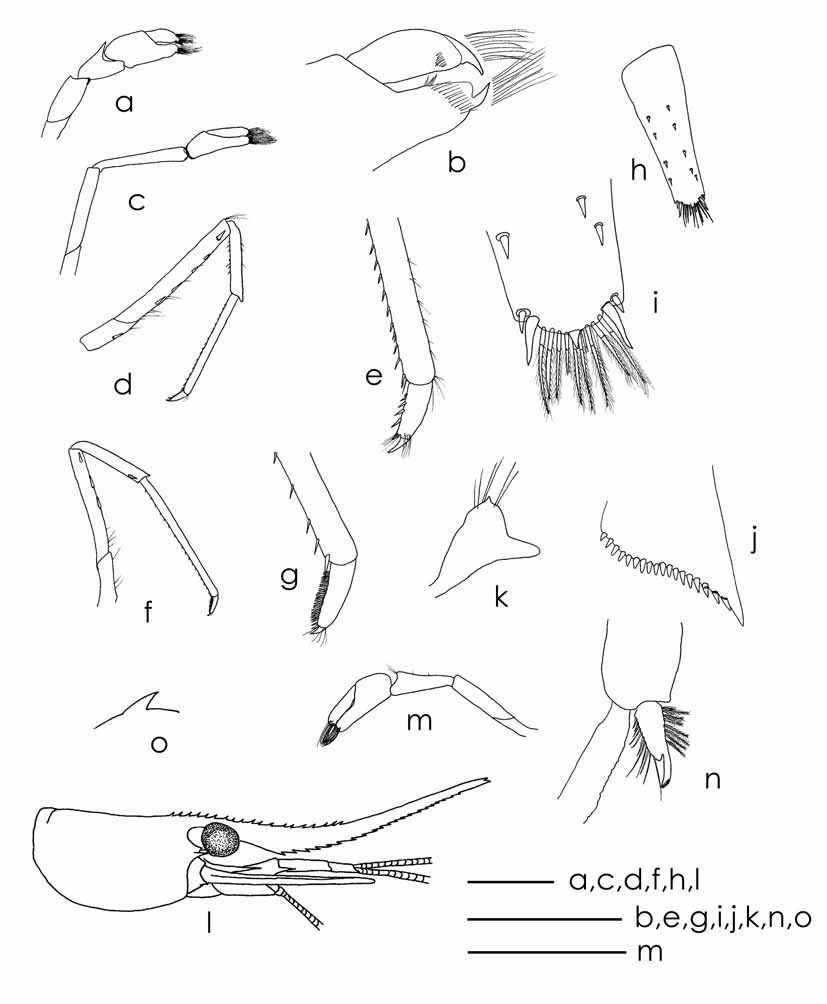
Description: Rostrum ( Fig.2 View FIGURE 2 a, d): long, 0.7–1.4 of cl, styliform, reaching to or overreaching distal end of scaphocerite, sigmoid, curved upward in distal half, tip slightly bent downwards, armed with 13–15 (14) teeth on dorsal margin, 3–4 of them situated on carapace behind orbital margin, distal 1/3 of dorsal margin unarmed, ventral margin with 4–7 teeth, distally wider spaced than proximally.
Eyes ( Fig.2 View FIGURE 2 a, d): well developed with cornea globular.
Carapace ( Fig.2 View FIGURE 2 a, d): Smooth, glabrous with sharp antennal spine placed at lower orbital angle or slightly below. Pterygostomial margin blunt but narrow, slightly produced forward, no teeth.
Antennule ( Fig 2 View FIGURE 2 k): Peduncle as long as carapace in male, about 0.7 cl in female, three-segmented; proximal segment as long as combination of second and distal segment, latter shorter than second; anterolateral process of proximal antennular segment blunt; all segments with submarginal plumose setae; stylocerite sharp, long, reaching to about half of second segment of peduncle.
Scaphocerite ( Fig.2 View FIGURE 2 l): Slightly longer than antennular peduncle, outer margin slightly concave, asetose, ending in a strong subapical spine; length about 3.5 times width, inner and anterior margins with long plumose setae.
Mouthparts as figured in Fig.2 View FIGURE 2 . Mandibels ( Fig.2 View FIGURE 2 e): Incisior process of mandibels ending in irregular teeth, molar process truncated. Upper lacinia of maxillule ( Fig. 2 View FIGURE 2 g) elongated with numerous small teeth on inner margin, palp slender with few plumose setae at tip, lower lacinia broadly rounded. Upper endites of maxilla ( Fig.2 View FIGURE 2 f) subdividet, palp fingerlike, scaphognatite tapering posteriorly with long hairs at distal end.
First maxilliped ( Fig.2 View FIGURE 2 h): with palp truncate ending in a short fingerlike tip.
Second maxilliped ( Fig. 2 View FIGURE 2 i) as typical for the genus.
Third maxilliped ( Fig.2 View FIGURE 2 j): With exopod, penultimate segment of endopod longer than proximal and distal segment.
First pereiopod ( Fig.3 View FIGURE 3 a): Stout, chela broader than second, about 1.9–2.3 times as long as wide, 1.3–1.7 times length of carpus; movable finger 1.3–2.3 times as long as wide, 0.4–0.7 times length of palm; tips of both fingers ( Fig.3 View FIGURE 3 b) with prominent claws; carpus deeply excavated disto-dorsally, 1.2–1.7 times as long as wide, 0.6–0.8 times as long as chela and about as long as merus.
Second pereiopod ( Fig.3 View FIGURE 3 c): More slender and longer than first pereiopod with chela 2.2–3.0 times as long as wide, 0.7–0.8 times length of carpus; movable finger 3.0–4.8 times as long as wide, 1.0–1.5 times length of palm, tips of fingers rounded; carpus slender 5.0–6.7 times as long as wide, not excavated distally, about as long as merus.
Third pereiopod ( Fig. 3 View FIGURE 3 d): Slender, dactylus ( Fig. 3 View FIGURE 3 e) 3.3–4.0 times as long as wide (terminal spine included) with 5–6 spines on flexor margin in addition to the terminal spine; propodus 10.1–14.0 times as long as wide, 3.9–5.3 times as long as dactylus; carpus 5.3–6.1 times as long as wide, 0.6–0.7 times as long as propodus, 0.5–0.6 times as long as merus; merus 7.6–9.2 times as long as wide, 1.7–2.1 times as long as carpus, bearing 5 strong, movable spines on posterior margin of outer surface.
Fifth pereiopod ( Fig. 3 View FIGURE 3 f): Dactyli ( Fig. 3 View FIGURE 3 g), 3.5–4.4 as long as wide with 26–29 spines on flexor margin; propodus slender, 12.8–20.5 times as long as wide distally, 4.5–6.2 times as long as dactylus; carpus 4.2–8.9 as long as wide, 0.5–0.6 times as long as propodus, 0.6–0.7 times as long as merus; merus 7.2–9.9 times as long as wide, bearing 3–4 strong, movable spines on posterior margin of outer surface.
Well developed epipods on first to fourth pereiopods.
First pleopod ( Fig.2 View FIGURE 2 b): Endopod elongated triangular in males, narrowing distally, 2.8 times as long as maximum wide, about 0.4 times as long as exopod, inner border with five marginal setae, outer margin with plumose setae, longer than those on inner margin, appendix interna well developed, arising from about ¼ distal part, clearly overreaching the tip of endopod; exopod 6.5 times as long as wide.
Second pleopod ( Fig.2 View FIGURE 2 c): Endopod in male about 0.8 times as long as exopod, appendix interna reaching about 2/3 of appendix masculina; appendix masculina slender, rod-shaped, reaching to about 0.6 times length of endopod, bearing a row of long spines on inner margin and distally.
Abdomen: Glabrous, sixth abdominal somite 0.49–0.56 times cl.
Preanal carina ( Fig.3 View FIGURE 3 k): High, shape as figured with an indistinct teeth.
Telson ( Fig.3 View FIGURE 3 h): 0.80–0.97 times cl in males 0.62–0.69 cl in females, longer than sixth abdominal segment, tapering posteriorly, ending in broadly convex margin bearing a median point above true posterior margin, dorsal surface with two rows of 4–6 movable spines including the pair at posterolateral angles; posterior margin ( Fig. 3 View FIGURE 3 i) with 7–12 setae, intermedial plumose setae much longer than lateral stout pair.
Uropodal diaeresis ( Fig.3 View FIGURE 3 j) with 17–18 spinules.
Egg size: 0.40–0.45 x 0.23–0.25 mm.
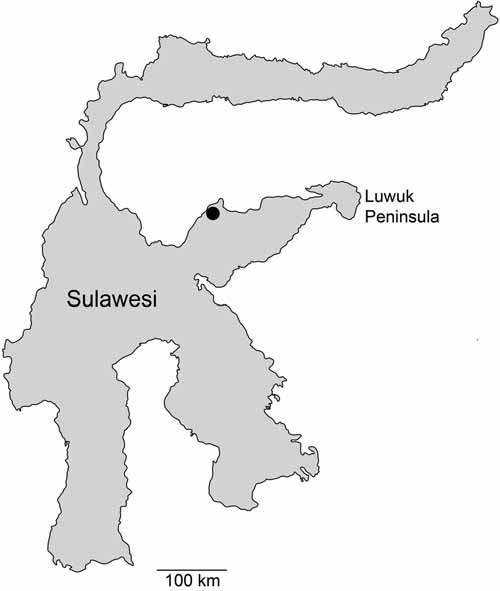
Habitat: All specimens were collected from a freshwater spring at the north coast of Luwuk Peninsula ( Fig. 1 View FIGURE 1 ), which is temporarily flooded with saltwater and represents a typical brackish water environment. The shrimp was predominantly found on rocky substrate.
Distribution: C. buehleri was original described by Roux from Bimoun, New Ireland, Papua New Guinea and is now reported from Sulawesi, Indonesia. The small and numerous eggs, indicative of a prolonged larval development, and the habitat (brackish water pool) suggest the possibility of a wider distribution range, though.
Remarks: C. buehleri was originally described by Roux in 1934 based on a single non-ovigerous specimen. Even though the description of Roux is brief and bears no information on the formation of the male sexual appendages and the egg size of ovigerous females, there is no doubt about the conspecificity of our specimens with C. buehleri . Our material differs from the measurements given by Roux only in the length to breadth ratio of the propodi of the 3rd (10.1–14.0 times as long as wide vs. 8 times reported by Roux) and 5th (12.8–20.5 times as long as wide vs. 8 times reported by Roux) pereiopods, but this might be merely an adaptation to local circumstances as discussed by Woltereck (1937a) for riverine and lacustrine populations of C. nilotica and should be treated as intraspecific variation. Furthermore the length to breadth ratios of these segments are very variable in the present specimens. The new material from Sulawesi, which includes both males and ovigerous females, provides further information on this species and its taxonomical position. The long, styliform rostrum, the long stylocerite and the long intermediate spines on the distal margin of the telson distinguish Caridina buehleri from all congeners described from the entire Indopacific region. With regards to the long stylocerite, which reaches to the middle of the second segment of the antennular peduncle, and the intermediate spines on the distal margin of the telson (longer than the lateral pair), the species mostly resembles C. serratirostris De Man, 1892 and C. celebensis De Man, 1892 . The following characters clearly differentiate C. buehleri from the two species: the form and denticulation of the rostrum with a long unarmed portion distally and the stouter first pereiopods with fingers much shorter than the palm (versus fingers longer than the palm in C. serratirostris and C. celebensis ), the carpus 1.2–1.7 times as long as wide, deeply excavated distally (vs. about four times as long as wide, not deeply excavated distally in C. serratirostris and C. celebensis ). Additionally C. buehleri possesses a long appendix interna on the male first pleopod which is lacking in C. serratirostris and C. celebensis ( De Man 1892; Chace 1997; Liang 2004; Cai & Shokita 2006). A long stylocerite reaching to the second segment of the antennular peduncle is also found in all members of the Caridina serrata species group (Cai 1999, Liang 2004). The long intermediate spines on the distal margin of the telson, the shape of the male first endopod and the small egg size, suggestive of a prolonged larval development, differentiate Caridina buehleri from all members of this species group, though. Moreover it seems unlikely that a member of this species group, comprising only species with large eggs and therefore presumably an abbreviated larval development which usually indicates a limited possibility for oceanic dispersal should be found disjunct on Sulawesi far from the known distribution range of these species in China. The prominent claws on the fingertips of the first pereiopods could possibly represent a specialized feeding adaptation of Caridina buehleri . While apical hooks or claws on fingers of atyid chelae were already discussed by Hickson (1888) in C. wyckii, Bouvier (1925) in his diagnosis of the genus Caridina and Gurney (1984) in her work on Madagascan atyid shrimps, none of them did venture to suggest a function for this character. C. buehleri was collected primarily on rocks and has a rather dark (brownish) body colouration. A second atyid shrimp, C. appendiculata , was found living in sympatry with C. buehleri but on a different substrate (macrophytes) and has a lighter and more conspicuous colouration. The fingers of this species have rounded tips like most other Caridina species. Clawed fingers might prove advantageous in feeding on small invertebrates like oligochaetes or juvenile insect larvae, while species without clawed finger tips are primarily feeding on detritus or single-celled periphyton ( Fryer 1960, Hussein & Obuid-Allah 1992). Analyses of the gut content of clawed-fingered species could be used to test this hypothesis.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |