
Mioptychopyge thebawi ( Reed, 1915 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5162.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:DD2279FA-E8F1-4951-A5CA-91082E875580 |
|
DOI |
https://doi.org/10.5281/zenodo.6816297 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB8784-6E42-1D11-FF3A-C1744B16F31D |
|
treatment provided by |
Plazi |
|
scientific name |
Mioptychopyge thebawi ( Reed, 1915 ) |
| status |
|
Mioptychopyge thebawi ( Reed, 1915)
Fig. 9 View FIGURE 9
1915 Ptychopyge thebawi sp. nov. Reed, p. 32–34, pl. 6, figs 5–8.
1915 Ptychopyge (Basilicus) titanica sp. nov. Reed, p. 35, pl. 6, figs 11–12, pl. 7, fig. 1.
1998 Mioptychopyge thebawi ( Reed, 1915) ; Zhou, Dean, Yuan & Zhou, p. 706.
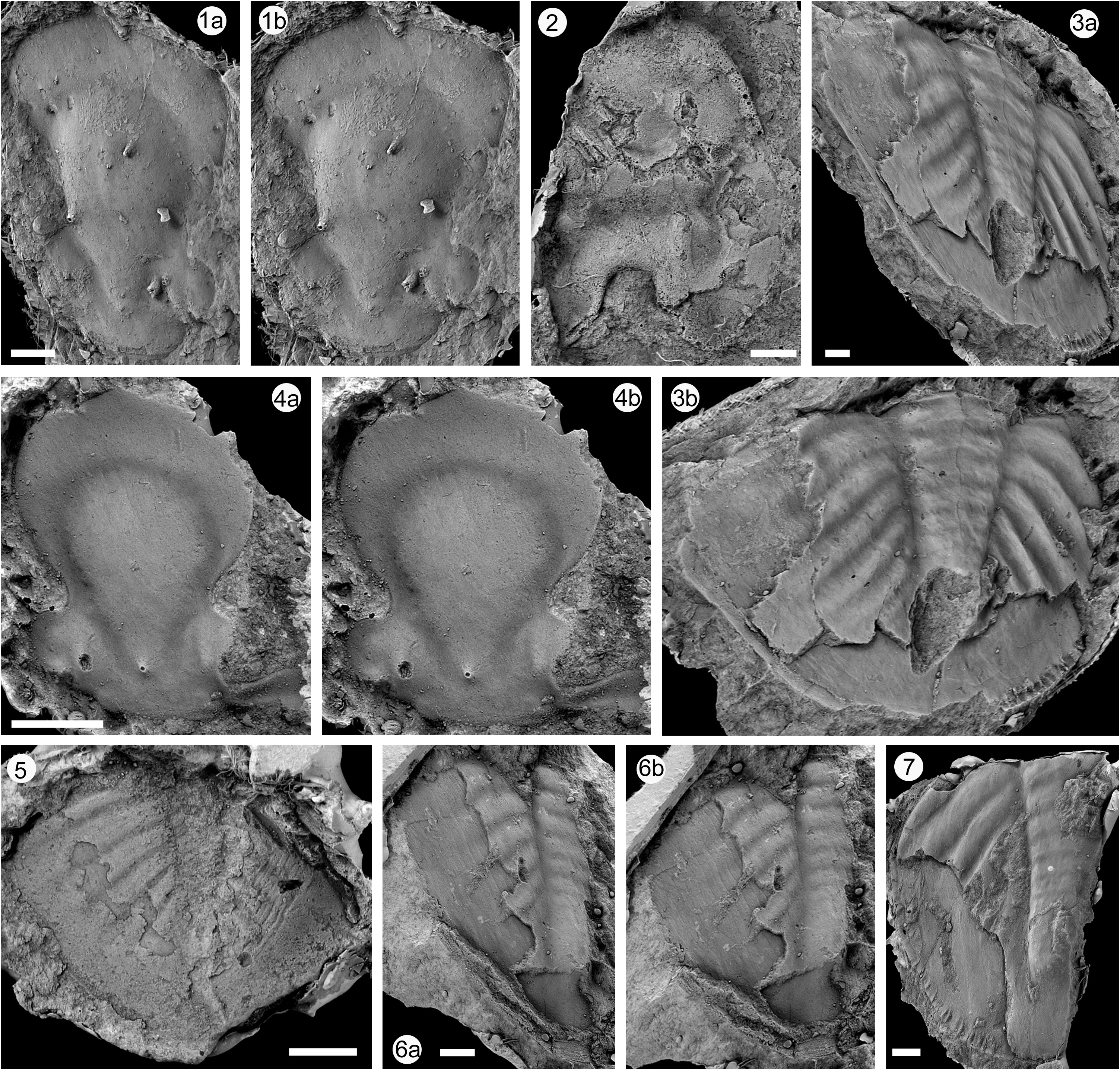
Material. Lectotype (selected here): cranidium from Hkawnhkok, Fig. 9.4 View FIGURE 9 ( Reed, 1915, pl. 6, fig. 7), GSI 11527. Hkawnhkok. Additional type material: cranidium from Nawng Yun, Fig. 9.1 View FIGURE 9 ( Reed, 1915, pl. 6, fig. 5), GSI 11525; cranidium from Mong Ha, Fig. 9.8 View FIGURE 9 ( Reed, 1915, pl. 6, fig. 6), GSI 11526; pygidium from Mong Ha, Fig. 9.5 View FIGURE 9 ( Reed, 1915, pl. 6, fig. 8), GSI 11528. Additional material: hypostome from Lilu, Fig. 9.3 View FIGURE 9 ( Reed, 1915, pl. 6, fig. 9), GSI 11529; 3 pygidia from Hwe Mawng, Figs 9.3 View FIGURE 9 , 6 View FIGURE 6 , 7 View FIGURE 7 ( Reed, 1915, pl. 7, fig. 1; pl. 6, figs 12, 11, 39 respectively), GSI 11533, 11532, 11531 respectively. All specimens from Hwe Mawng Beds (uppermost Katian) at Hkawnhkok, Hwe Mawng, Lilu, Mong Ha, Nawng Yun.
Description. Most of the material illustrated by Reed (1915) is a little distorted, and plausible retrodeformations are also used herein. Cranidia are not transversely convex and would originally have been of similar width (tr.) along the posterior margin as they are long sagitally. Glabella occupies about 70% cranidial length (sag.), with maximum width across the frontal lobe close to 60% sag. length. Frontal lobe pyriform and extended (sag.), with anterior part in front of maximum width forming a semicircle in outline, and posterior part tapering regularly to level of glabellar tubercle; furrows defining it rather uniformly shallow. Although the furrows appear continuous the posterior parts are slightly deeper and here comprise the S1 glabellar furrow. The probable course of the posterior part of the axial furrow is shown on the right-hand side of the holotype as a shallow furrow running more or less exsagitally. Hence the glabella is constricted at the level of the eyes to about two-thirds its maximum width at the front, and then widens posteriorly to a similar transverse width at the posterior margin. Prominent glabellar tubercle at one-fifth glabella length, opposite posterior part of the palpebral lobes, which are highly curved with weak rims, and of length (exsag.) one quarter that of glabella. Although they are close to the glabella there are small (exsag.) inflated areas adjacent to the palpebral lobes, which lie outside the axis. Postocular fixed cheek is very narrow (exsag.) proximally, wider laterally, and much of it is taken up by wide posterior border furrow that widens laterally. Preocular fixed cheeks merge with wider, flat preglabellar area. Facial sutures are recurved forwards behind the eyes, anterior branches diverging at about 40 degrees to sag. Line before curving adaxially in a smooth curve to meet at midline in a distinct point, so there presumably supramarginal. Free cheek not assigned. A hypostome ( Fig. 9.2 View FIGURE 9 ) is imperfectly preserved but shows a long (sag.) middle body that is compatible with the long anterior glabellar lobe of this species. The relatively short fork has a U-shaped profile, and the lateral border is not flared, as in several other asaphid genera.
A pygidium ( Fig. 9.5 View FIGURE 9 ) assigned by Reed (1915) is incomplete but apparently not distorted, and of a size that would be appropriate for the lectotype cranidium; it would have been two-thirds as long as wide when complete. Five pleural ribs, of which four are well defined, the fifth fainter, while the pleural furrows fade out over half-way across the pleural field. The pygidial axis is not well-preserved, but must have had narrow proportions (approximately three times as long as wide), and presumably a minimum of five axial rings to accord with the pleural ribs. The doublure, closely reflexed against the dorsal surface and carrying terrace ridges, extended quite far beneath the distal parts of the pleural furrows, but did not extend to the axis. Reed (1915) also described some very large pygidia under the name Ptychopyge (Basilicus) titanica , all of which have undergone distortion. When retrodeformed ( Figs 9.3b View FIGURE 9 , 6b View FIGURE 6 ) these are plausible as large specimens of M. thebawi , showing five ribs which fade towards the pygidial margin and a similarly narrow pygidial axis. A paradoublural line on the largest pygidium indicates that the doublure closely approached, but did not reach, the axis. A sixth axial ring is observed and long terminal piece apparently without dorsal furrows. On the small pygidium there are faint indications of interpleural furrows, but their apparent absence in the large pygidia is probably accounted for by changes during later ontogeny.
Discussion. Zhou et al. (1998) assigned Ptychopyge thebawi Reed, 1915 , to their new genus Mioptychopyge , when redescribing the type species, M. trinodosus Zhang, 1981 , from Yunnan Province. These authors assigned an additional three Chinese species to Mioptychopyge , all of them named later than M. thebawi . Mioptychopyge trinodosus has a distinct anterior border furrow on the cranidium, and a wider pygidial axis than M. thebawi , but shorter and weaker pygidial pleural ribs. The comparison with the typically Baltic genus Ptychopyge and its allies is based on the ogival form of the anterior branches of the facial sutures, which must have been supramarginal. However, it is also possible to compare Asian species with the Subfamily Birmanitinae , as discussed above, for example in the elongate pyriform glabella with prominent posterior tubercle. Many Baltic ptychopygines have a pair of distinctive inflated nodes at the anterior proximal edges of the fixigenae which are not seen in the Chinese species (see discussion in Turvey, 2007, p. 361), and this may prove to be a synapomorphy of the former group. Birmanites brevicus Xiang & Zhou, 1987 from Tarim, revised by Zhou et al. (1998), is very similar to M. thebawi with the exception of a rounded anterior junction of the facial sutures at the cephalic margin. Its pygidium does not have the curved ribs typical of Birmanites . It may prove to be closer to Mioptychopyge . Mioptychopyge suni ( Endo, 1935) revised by Turvey (2007) is also generally similar to M. thebawi , but has much larger palpebral lobes and feebly developed pygidial pleural ribs. Closer to M. thebawi is a species referred by Turvey (2007) to Opsimasaphus , O. pseudodawanicus ( Lu, 1975) , from the Siliangssu Formation, southern China. Although the palpebral lobes on the well-preserved cranidium illustrated by Turvey (2007, pl. 3, fig. 14) are larger than on Reed’s M. thebawi , this cranidium is also much smaller, and it is generally true in asaphids that the proportional length of the eye diminishes during later ontogeny (e.g. Asaphellus see Fortey & Owens, 1991). It is possible that this difference is just a matter of ontogeny, and in other features the cranidia are similar (compare retrodeformed Fig. 9.4b View FIGURE 9 herein with Turvey, 2007, pl. 3, fig. 13) apart from a more posteriorly curved anterior branch of the facial suture in O. pseudodawanicus , and a fragmentary larger cranidium ( Turvey, 2007, pl. 4, fig. 1) does appear to show a wider preglabellar area. The pygidium of O. pseudodawanicus is like that of M. thebawi in having strongly developed pleural ribs, but up to eight are developed on the former. Turvey (2007) gave a lengthy discussion on the generic placement of pseudodawanicus , and the possible differentiating features of Opsimasaphus , and clearly recognised that Birmanites , Opsimasaphus and Mioptychopyge comprise a closely related, and indeed intergrading plexus. The incomplete information we have on M. thebawi does not help arbitration on these genera, and it is here retained in Mioptychopyge as Zhou et al. (1998) suggested.

| GSI |
Geological Survey of India |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Birmanitinae |
|
Genus |