
Wirada mexicana, Campuzano & Ibarra-Núñez, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4457.3.13 |
|
publication LSID |
lsid:zoobank.org:pub:BBA4B95A-E83B-4A91-9948-7FB12A742E16 |
|
DOI |
https://doi.org/10.5281/zenodo.5955704 |
|
persistent identifier |
https://treatment.plazi.org/id/FD281C8C-FEA2-4786-BDCB-DED0655C3D6A |
|
taxon LSID |
lsid:zoobank.org:act:FD281C8C-FEA2-4786-BDCB-DED0655C3D6A |
|
treatment provided by |
Plazi |
|
scientific name |
Wirada mexicana |
| status |
sp. nov. |
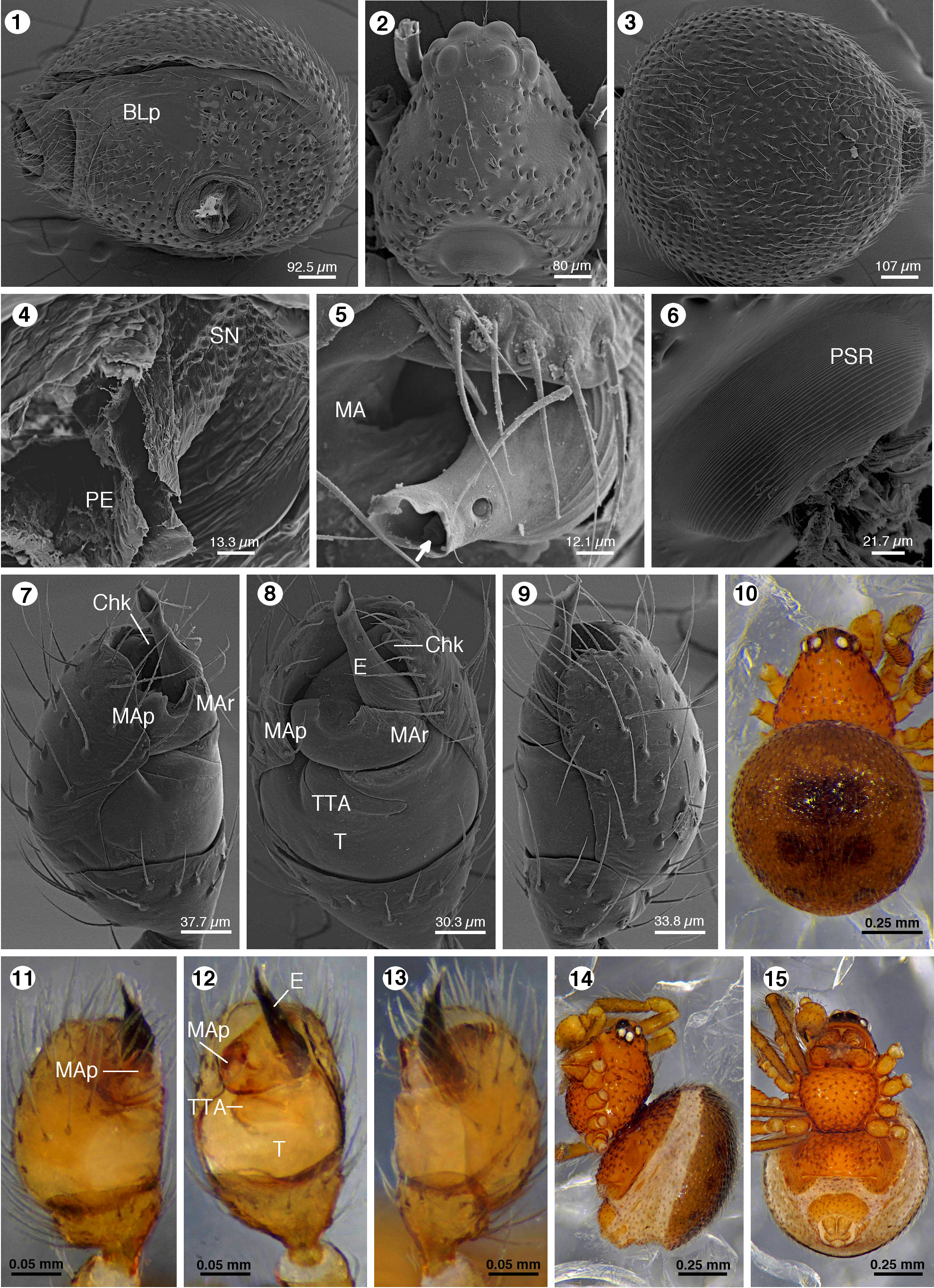
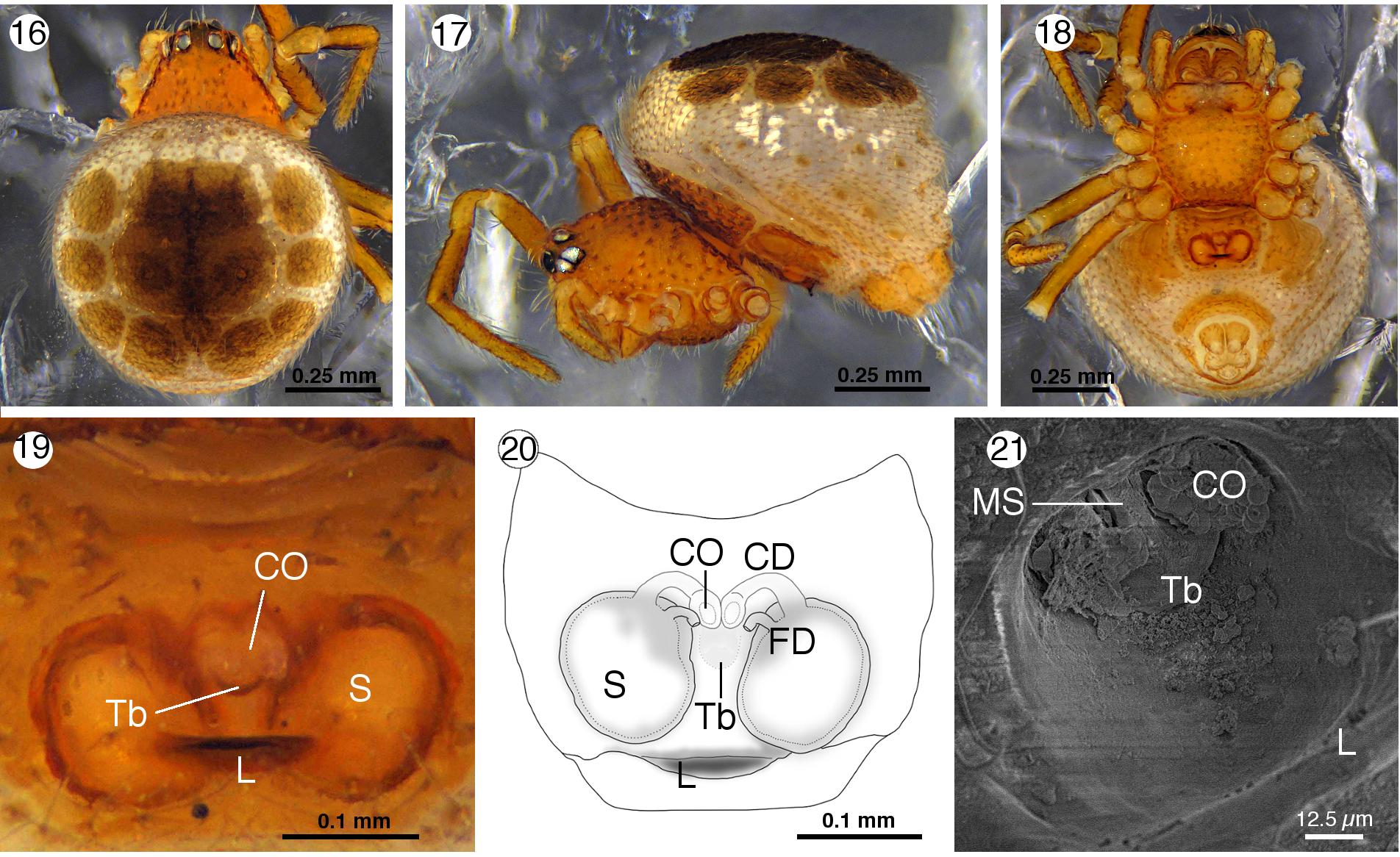
Wirada mexicana View in CoL new species ( Figs 1–22 View FIGURES 1–15 View FIGURES 16–21 View FIGURE 22 )
Type material. Holotype male: MEXICO : Chiapas: Municipio de Motozintla, Cerro Boquerón (15°13’55.1’’ N, 92°18’21.6’’W, 2332 m), April 29, 2015, on understory of tropical mountain cloud forest, beating tray, E. Campuzano, E. Chamé, L. Gallegos, S. Moreno & G. Sánchez (ECOTAAR-9010) GoogleMaps . Allotype female: Same data, except March 25, 2015, searching, E. Campuzano, L. Gallegos, H. Montaño & S. Moreno (ECOTAAR-9009) . Paratypes: 1 ♂, same data as holotype, except searching ( AMNH) ; 1 ♂, same data except February 25, 2015, netting, E. Campuzano, H. Montaño & S. Moreno ( CAS) . 1 ♂, La Sepultura Biosphere Reserve, Municipio de Villacorzo, Cerro Bola (16°08’20.7’’ N, 93°36’36.4’’ W, 1769 m), August 3, 2016, on understory of tropical mountain cloud forest, netting, E. Campuzano, H. Montaño, S. Moreno & E. Chamé (ECOTAAR-9015) GoogleMaps . 1 ♂, same data, except beating (ECOTAAR-9016) GoogleMaps . 1 ♂, Tacaná volcano Biosphere Reserve, Municipio de Unión Juárez , Talquián (15°05’38’’ N, 92°06’06’’ W, 2044 m), March 5, 2009, on understory of disturbed tropical mountain cloud forest, searching, J. Maya, G. Ibarra, J. A. López & E. Senties (ECOTAAR-6476) GoogleMaps . 1 ♂, San Luis Potosí: Municipio de Xilitla, Las Pozas (21°23’50’’ N, 98°59’38’’ W, 600 m), June 10–15, 2012, on understory of tropical wet forest fragment, beating tray, Aracnolab leg ( CNAN) GoogleMaps .
Etymology. The specific epithet refers to the country of origin.
Diagnosis. Males of this species differ from other Wirada species by having a straight embolus ( Figs 7–9, 11–13 View FIGURES 1–15 ), all other species have a coiled embolus ( Levi 1963, figs 14, 19, 20; Levi 1967 fig. 30; Lise et al. 2009 figs 6, 8, 29, 32). They differ also because the tip of the prolateral projection of the median apophysis is truncated and does not surpass the distal border of the cymbium ( Figs 7–9 View FIGURES 1–15 ), whereas in all other species the prolateral projection of the median apophysis is tapered and surpass the distal border of the cymbium ( Levi 1963, figs 14, 19, 20; Levi 1967, fig 31; Lise et al. 2009, figs 6, 7, 29, 30,31). The female differs from the species with known females by the copulatory openings separated from labium of epigynum by about two times their diameter (at level of the anterior border of the spermathecae, Figs 19–21 View FIGURES 16–21 ), the other species have the copulatory openings separated from labium of epigynum by one times their diameter or less ( Levi 1963, figs 16, 22; Lise et al. 2009, figs 24, 42). In W. mexicana n. sp. the copulatory ducts are short and laterally directed to enter the spermathecae at their anterior border ( Fig. 20 View FIGURES 16–21 ), while in the other species the copulatory ducts are longer and located between the two spermathecae ( Levi 1963, figs 15, 21; Lise et al. 2009, figs 18, 38).
Description. Male holotype. Color: carapace, labium, endites, sternum and chelicerae red-fulvous, concolorous ( Figs 10, 14–15 View FIGURES 1–15 ). Dorsal opisthosomal scutum chestnut, with the sigilla surrounded by darker circular spots, four centrals bigger than laterals ( Fig. 10 View FIGURES 1–15 ). Ventral scutum as carapace ( Figs 14–15 View FIGURES 1–15 ). Lateral areas light brown, with the sigilla fulvous ( Fig. 14 View FIGURES 1–15 ). Spinnerets and ring around them fulvous ( Fig. 15 View FIGURES 1–15 ). Legs fulvous darker on the distal segments. Palp light fulvous, with embolus black ( Figs 11–13 View FIGURES 1–15 ). Habitus: Total length 1.55. Carapace 0.61 long, 0.52 width, with chitinous setiferous bridges arranged longitudinally in the middle and diagonally on lateral areas ( Figs 2, 10 View FIGURES 1–15 ). Carapace with prosomal stridulatory ridges on its posterior border ( Fig. 6 View FIGURES 1–15 ). Ocular area with a few simple setae ( Fig. 2 View FIGURES 1–15 ). Eye sizes and interdistances: AME 0.06, ALE 0.046, PME 0.052, PLE 0.052, AME-AME 0.017, AME-ALE 0.009, PME-PME 0.06, PME-PLE 0.017. AER and PER almost straight ( Figs 2, 14–15 View FIGURES 1–15 ). Clypeus 0.20 high. Cheliceral promargin with a bicuspid tooth. Endites 0.21 long, 0.13 width. Labium 0.06 long, 0.17 width. Sternum 0.33 long, 0.39 width, convex, with chitinous setiferous bridges, intercoxal sclerites present, posterior edge nearly straight, extended beyond of coxae IV, about half as wide as anterior border ( Fig. 15 View FIGURES 1–15 ). Opisthosoma 0.94 long, 0.85 width, circular in dorsal view. Dorsal scutum microsculptured with backward directed setae ( Fig. 3 View FIGURES 1–15 ) and twelve conspicuous circular sigilla, four at central area and eight around these ( Figs 3, 10 View FIGURES 1–15 ). Ventral scutum surrounding pedicel and covering epigastrium with chitinous setiferous bridges, book lung plates visible ( Figs 1, 14–15 View FIGURES 1–15 ); stridulatory nubbins at pedicel base ( Fig. 4 View FIGURES 1–15 ); spinnerets surrounded by four sclerotized plates, anterior and posterior bigger than laterals, forming a ring ( Fig. 14–15 View FIGURES 1–15 ). Membranous area between ventral and dorsal scuta with sputtered oval sigilla and sclerotized setal bases ( Fig. 14 View FIGURES 1–15 ). Legs measurements: Femora I 0.45, II 0.43, III 0.37, IV 0.44. Patella I 0.17, II 0.17, III 0.15, IV 0.19. Tibiae I 0.27, II 0.26, III 0.24, IV 0.28. Metatarsi I 0.25, II 0.23, III 0.22, IV 0.24. Tarsi I 0.26, II 0.26, III 0.23, IV 0.28. Palp: Tibia goblet-shaped ( Figs. 8, 12 View FIGURES 1–15 ), cymbium oval, convex ( Fig. 9, 13 View FIGURES 1–15 ); tegulum convex occupying the proximal two thirds of bulb ( Figs. 7–9 View FIGURES 1–15 ); theridioid tegular apophysis embedded in the tegulum, spreading to less of distal half (between tibia and median apophysis) ( Figs. 8, 12 View FIGURES 1–15 ). Median apophysis distal to tegulum, transverse, u-shaped, partially encircling the embolus base to reach the cymbial hook dorso-ectal to embolus base ( Figs 5, 7–8 View FIGURES 1–15 ); tip of prolateral process of median apophysis truncated, not surpassing the cymbium distal border, lightly sclerotized ( Figs 7, 12 View FIGURES 1–15 ); retrolateral process of median apophysis inconspicuous, crest-shaped, hyaline ( Figs 8, 12 View FIGURES 1–15 ). Embolus straight and cylindrical, its length about three and a half times as wide at its base, its tip opening oblique, partially hyaline and with a small orifice near its tip ( Figs 5, 7–9, 11–13 View FIGURES 1–15 ).
Female allotype. Color: as holotype, except dorsal scutum, pallid yellowish with chestnut spots on sigilla ( Figs 16–17 View FIGURES 16–21 ). Habitus: Total length 1.62. Carapace 0.64 long, 0.52 width, with chitinous setiferous bridges arranged as holotype. Ocular area with few simple setae ( Fig. 16 View FIGURES 16–21 ). Eye sizes and interdistances: AME 0.07, ALE: 0.043, PME 0.052, PLE 0.052, AME-AME 0.013, AME-ALE 0.009, PME-PME 0.06, PME-PLE 0.023. AER and PER almost straight ( Figs 16–17 View FIGURES 16–21 ). Clypeus 0.11 high. Cheliceral promargin with a bicuspid tooth. Endites 0.20 long, 0.12 wide. Labium 0.08 long, 0.16 width. Sternum 0.31 long, 0.41 width, shape and texture as holotype ( Fig. 18 View FIGURES 16–21 ). Opisthosoma 0.98 long, 0.98 width, shape and structure as holotype ( Fig. 16 View FIGURES 16–21 ). Sclerotized ring of spinnerets as holotype, except anterior plate narrower. Dorsal scutum with fourteen conspicuous circular sigilla, four at central area and ten around these ( Fig. 16 View FIGURES 16–21 ). Ventral scutum divided in four plates ( Figs 17–18 View FIGURES 16–21 ), one dorsal to pedicel, covered with chitinous setiferous bridges ( Fig. 17 View FIGURES 16–21 ), tree others on the epigastrium, one surrounding the epigynal plate and two laterals encircling the book lungs plates ( Figs 17–18 View FIGURES 16–21 ). Legs measurements: Femur I 0.51, II 0.46, III 0.37, IV 0.45. Patella I 0.17, II 0.17, III 0.17, IV 0.17. Tibia I 0.27, II 0.24, III 0.21, IV 0.33. Metatarsi I 0.21, II 0.15, III 0.17, IV 0.23. Tarsi I 0.29, II 0.19, III 0.23, IV 0.29. Epigynum with a sclerotized labium projected ventrally from posterior border of atrium ( Figs 17–21 View FIGURES 16–21 ). Copulatory openings separated from labium of epigynum by about two times their diameter, separated by a septum that has a rear transverse bar ( Figs 19, 21 View FIGURES 16–21 ). Copulatory ducts short, laterally directed, forming a short arc before enter the spermathecae at their anterior border and slightly anterior to fertilization ducts insertions.
Variation. Males (n = 12): total length 1.1–1.3; carapace length 0.61–0.68; carapace width 0.52–0.6; abdomen length 0.9–1.05; abdomen width 0.75–0.91; femur IV 0.41–0.5; patella I 0.17–0.2; IV 0.18–0.23; tibia I 0.22–0.33; IV 0.28–0.35. Live specimens have chestnut color in both sexes. In preserved specimens color is degraded from chestnut to fulvous or yellowish.
Additional material examined. 1♂, MEXICO : Chiapas : El Triunfo Biosphere Reserve, core area I, Municipio de Mapastepec (15°39’48.4’’ N, 92°48’18’’ W, 2082 m), March 18, 2014, on understory of tropical mountain cloud forest, netting, E. Campuzano, J. Gómez, H. Montaño & G. Angulo (ECOTAAR-9007) GoogleMaps . 1♂, La Sepultura Biosphere Reserve, Muncipio de Villacorzo, Cerro Bola (16°08’20.7’’ N, 93°36’36.4’’ W, 1769 m), March 30, 2016; on understory of tropical mountain cloud forest, searching, E. Campuzano, H. Montaño, S. Moreno & E. Chamé (ECOTAAR-9012) GoogleMaps . 1♂, same data except April 26, 2016 (ECOTAAR-9013) GoogleMaps . 1♂, same data except August 3, 2016 (ECOTAAR-9017) GoogleMaps . 1♂, same data except July 5, 2016, beating tray (ECOTAAR-9014). GoogleMaps
Distribution. Mexico: Chiapas and San Luis Potosí ( Fig. 22 View FIGURE 22 ).
Natural history and habitat. Most of the studied specimens were collected on the understory of cloud forests from southern Mexico at elevations higher than 1700 m. However, the most northern record is from a tropical wet forest fragment at 600 m ( Fig. 22 View FIGURE 22 ), showing that the distribution range of this species likely comprises a greater number of habitats (and elevations) than here reported. A similar distribution pattern was observed in W. punctata ( Levi 1963) . Nevertheless, the species of Wirada are rare in the sense they are not easily found. Concerning the biology, Figure 21 View FIGURES 16–21 shows the CO obstructed by organic material, and Figure 5 View FIGURES 1–15 shows what could be solidified seminal fluid inside the embolus tip, suggesting the ability to produce a mating plug, as in other theridiid species ( Knoflach 2004).
Taxonomic notes. Levi & Levi (1962) and Levi (1963) referred as “radix” and “medium apophysis” what we homologize respectively to median apophysis and theridioid tegular apophysis (both following Agnarsson 2004). Levi and Levi (1962) noted for Wirada “Palp with paracymbial hook functional”, and “…the large triangular sclerite that fits into the paracymbial hook is the radix. It would be the only genus in which the radix fits against the paracymbial hook.” We followed the criteria and terminology of Agnarsson et al. (2007): “the median apophysis (MA) is often concealed in back of the bulb in the unexpanded palp, forms part of the uniquely theridiid BC[bulb-cymbium]-lock mechanism…” and “The TTA may be embedded within the T but is never fused to it.” Our images show clearly that the MA is interacting with the Chk to lock the bulb in the palp, and part of it is in the back of the bulb ( Figs 5, 7–8 View FIGURES 1–15 ), and also it is clearly visible that the TTA is embedded within the T for each species in this genus. Wirada mexicana n. sp. shows notable differences on the genital structures regarding its South American congeners and is the first North American genus record. Therefore, we hypothesize that Wirada mexicana n. sp. could correspond to a different subgenus. Such idea also could be supported by the seeming absence of the genus in Central America. However, it is not clear if the current geographic distribution of the species in the genus is a consequence of a vicariant distribution, or a deficiency of sampling the suitable habitats, as suggested by Lise et al. (2009). Additionally, the morphology of the palps of W. tijuca and W. araucaria differs only in minor details, and both species were found in Brazil (near the Atlantic coast). Since some Wirada species have a broad distributional range, it is possible that W. araucaria could be a junior synonym of W. tijuca , but verify this requires to compare the respective types.
Agnarsson (2004) suggested the possible inclusion of Wirada in the subfamily Pholcommatinae based in the synapomorphies of this group. Nevertheless, Wirada lacks most of these synapomorphies, sharing only cymbial hook on ectal margin, and embolus base shifted ectally and partially hidden by cymbium. Thus, with the information to date, it is no clear to which theridiid group belongs Wirada . It will be necessary to collect more specimens in both subcontinents and to redescribe some species to improve the knowledge of this genus.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |