
Pleurothallis carpishensis Ocupa, M.M.Jiménez & Mark Wilson, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.629.2.6 |
|
DOI |
https://doi.org/10.5281/zenodo.10390966 |
|
persistent identifier |
https://treatment.plazi.org/id/03C78782-FFA8-FFA7-6085-E94EFBD8FDDB |
|
treatment provided by |
Plazi |
|
scientific name |
Pleurothallis carpishensis Ocupa, M.M.Jiménez & Mark Wilson |
| status |
sp. nov. |
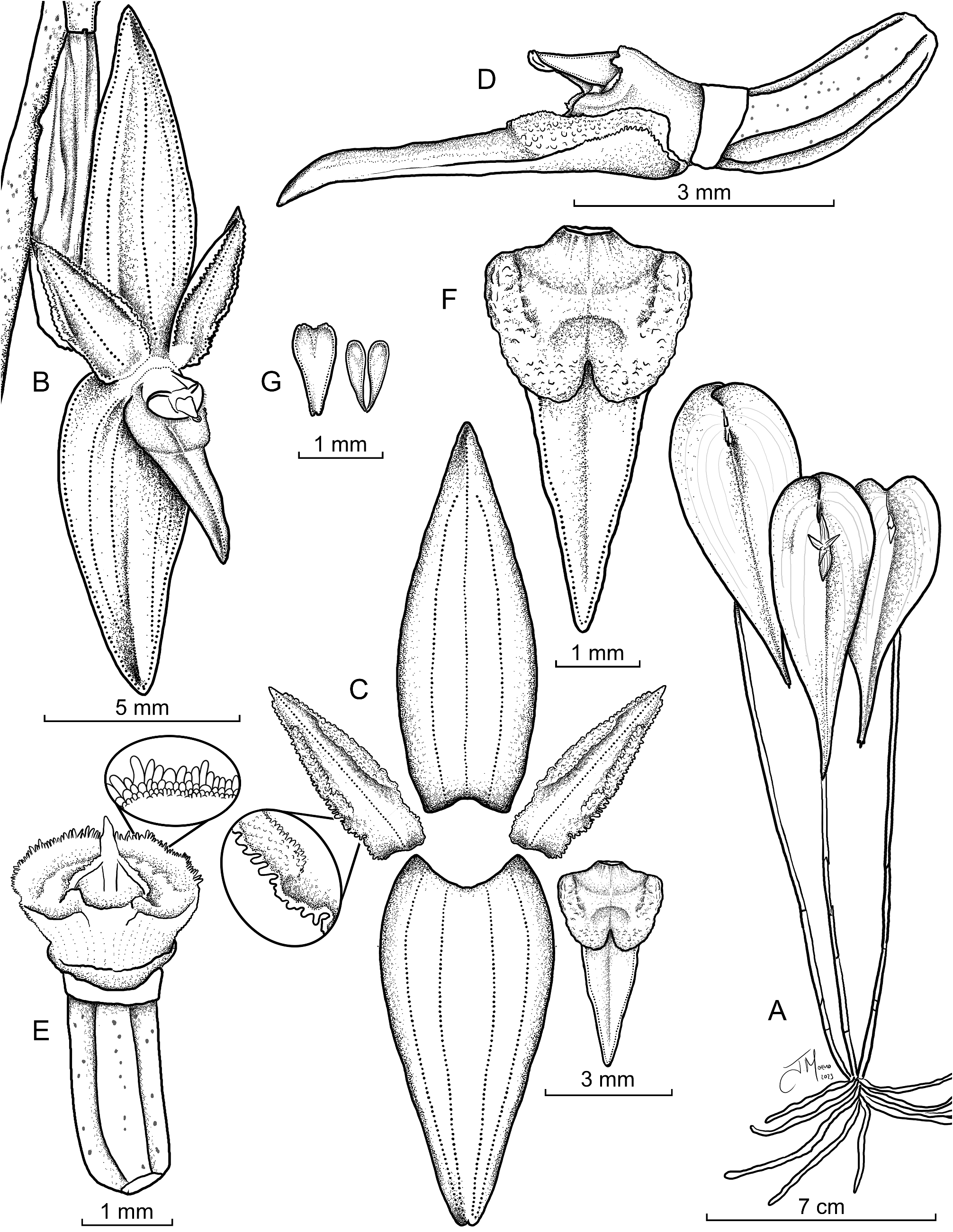
Pleurothallis carpishensis Ocupa, M.M.Jiménez & Mark Wilson View in CoL , sp. nov. ( Figures 2–3 View FIGURE 2 View FIGURE 3 )
Type: — PERU. Huánuco: Prov. Huánuco, Distrito Chinchao , Área de Conservación Regional Bosque Montano de Carpish, cerca del caserío Santa Rosa de Quives , 1722 m, 22 March 2022, L. Ocupa 301 (holotype: USM!) .
Morphologically similar to P. radula but is distinguished by the inflorescence that arises from a depressed, oblong, obtuse spathaceous bract (vs. erect, oblanceolate and sub-truncate), dorsal sepal lanceolate (vs. ovate to ovate-elliptic), petals oblong-lanceolate, apiculate, with large papillae on the margins (vs. narrowly lanceolate, acute and serrate), a triangular, sub-acute lip (vs. ovate to oblong-ovate, acute), with a callus broadly ovate and emarginate, papillose-verrucose (vs. callus glabrous, 1 mm long) and column edge surrounding stigmatic surfaces ciliate (vs. slightly papillate).
Description:— Epiphytic herb, caespitose, up to 23 cm tall, roots ca. 1.0 mm in diameter, slender, flexuous. Ramicauls suberect to erect, terete, ca. 8.0–20.0 cm long, 0.9–1.1 mm in diameter, curved at the apex, forming an angle of about 90° in the abscission layer, enclosed by two papyraceous, sulcate, pale brown, tubular sheaths, upper sheath ca. 20.0–23.0 × 3.0 mm, basal sheath ca. 8.0–12.0 × 3.0 mm, unifoliate. Leaves borne at the apex of the ramicaul, pale green, thinly coriaceous, deflexed toward the stem, concave, narrowly ovate, apically somewhat falcate, with a navicular concavity at the base of the spathe, margins involute, acuminate, tridentate sometimes with a longer central tooth, ca. 5.2–9.3 × 2.9–3.7 cm, base sessile, cordate, lobes unequal, sometimes overlapping. Inflorescence a solitary flower, resting on a navicular concavity at the base of the bract and borne from a depressed, conduplicate, oblong, obtuse spathaceous bract at the base of the leaf, ca. 8.5–9.5 mm long, striate, yellowish-green, becoming light brown, dry-papyraceous when mature, dotted with black, concealing peduncle, pedicel and floral bract; peduncle terete, up to 3 mm long; floral bracts infundibuliform, membranaceous, 3.5–5.0 mm long; pedicel terete, ca. 4.0–5.0 mm long; ovary terete, pale green, somewhat incurved, 6-furrowed, ca. 2.7–3.0 mm long, with black dots all over the surface. Flowers resupinate, spreading, yellow-hyaline, column white, anther cap cream, pollinia yellow; dorsal sepal erect, slightly concave, lanceolate, sub-acute, 8.0–10.3 × 3.2–3.8 mm, 3-veined; lateral sepals connate into a reflexed, ovate-lanceolate synsepal, centrally concave-channeled, obtuse, minutely emarginate, 8.0–10.0 × 4.5–4.8 mm, each half 2-veined; petals porrect, shallowly concave, oblong-lanceolate, apiculate, thickened apex, with large papillae on the margins, 4.5–6.0 × 1.5–2.0 mm, single-veined, abaxially 1-keeled; lip thick, fleshy, triangular, channeled in the midline, sub-acute, apex sometimes slightly recurved, 4.5–5.0 × 2.0– 2.5 mm, with three incomplete nerves, abaxial side foveolate, adaxial side cellular-papillose, without glenion; callus basal, widely ovate, emarginate, occupying 1/3 of the lip length, waxy, papillose-verrucose, the papillae irregularly coarse, especially at the margins, 1.7 × 2.3 mm, with two oblique, lateral, marginal and elevated keels; column short, stout, terete, ca. 1.0 × 2.0 mm, periphery of stigmatic surfaces ciliate, rostellum apical, rostellar flap long, linear, obtuse; anther apical, incumbent, papillose at the margins, anther cap cucullate, long-ovate, sub-cordate, 2-celled, 1.2 × 0.5 mm; stigma apical, reniform; pollinia 2, narrowly obovoid, complanate, 0.8 mm long, brought together by a minute viscidium.
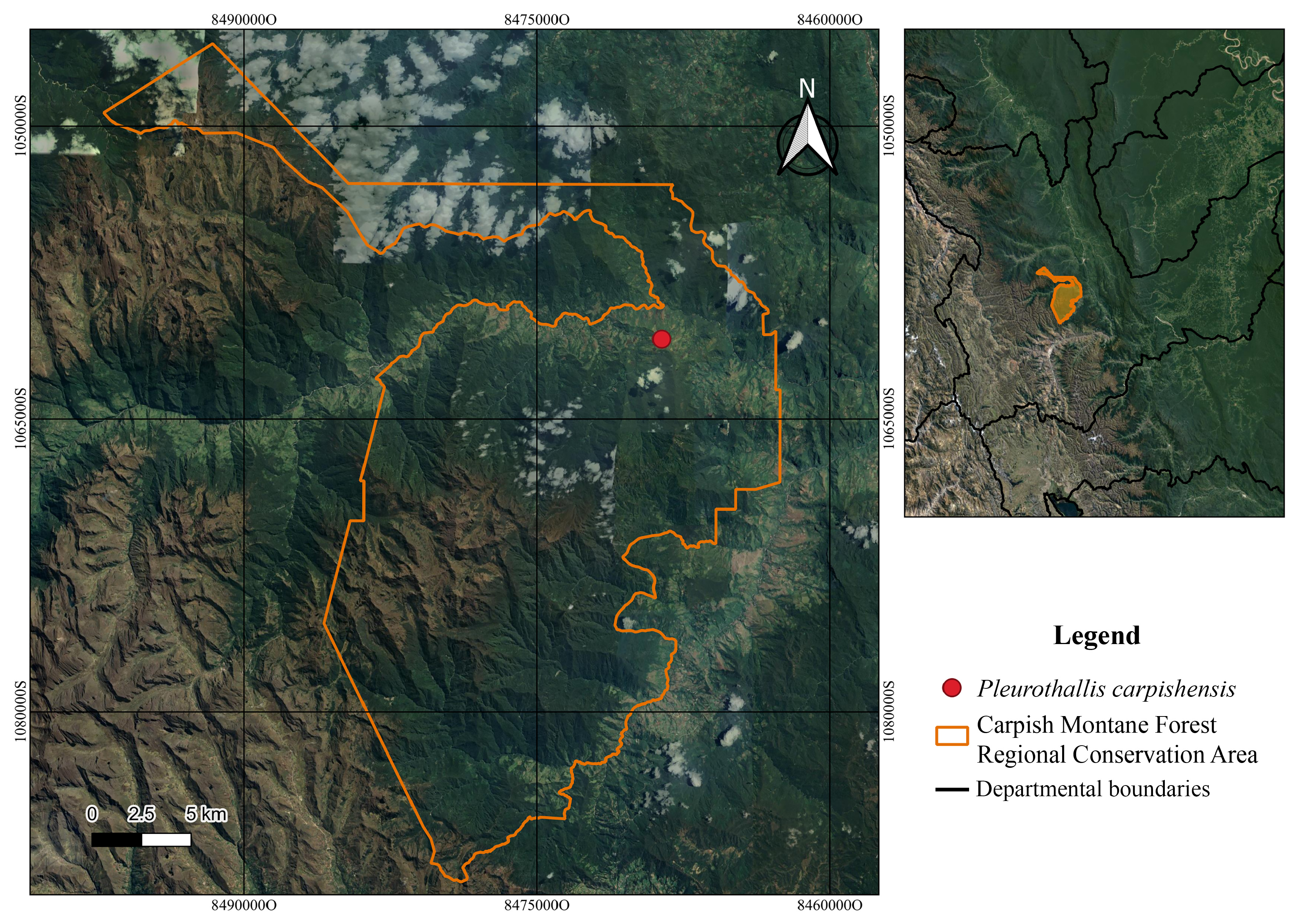
Distribution and ecology: — Pleurothallis carpishensis is known only from a small area on the eastern slope side of the central branch of the Peruvian Andes, forming part of the ecosystem known as the Yunga montane forest ( MINAM 2018, 2019), between 1720 and 1730 m elevation. This locality corresponds to the Carpish Montane Forest, located in the district of Chinchao, in the department of Huánuco ( Figure 4 View FIGURE 4 ). Individuals of this species grow as epiphytes on branches located in the tree canopy, especially in the zone IV ( Johansson 1974), at ~ 8 m above the ground ( Figure 5 View FIGURE 5 ). The flowering of this species occurs, at the beginning of the rainy season, between March and April, and during the dry season, between August and September.
Etymology: —In reference to the type locality, the Carpish Montane Forest Regional Conservation Area, Huánuco, Peru, where this species was discovered.
Conservation status: —This species is only known from the type collection in central Peru. The status of the population present in the Carpish Montane Forest is unknown; however, the habitat of this species may be affected by anthropogenic activities, such as deforestation and burning of trees, which occur in the area ( Figure 6 View FIGURE 6 ). Until a more comprehensive assessment can be made, the species should be considered "Data Deficient" ( DD) according to the IUCN criteria ( IUCN 2019).
Taxonomic Discussion: — Pleurothallis carpishensis can be distinguished by the pale yellow-to-hyaline flowers, that are borne from a depressed spathaceous bract that lies over a navicular concavity on the leaf, by petals oblong-lanceolate, apiculate, with large papillae on the margins and a triangular, sub-acute lip, with a callus basal, widely ovate, emarginate, waxy, papillose-verrucose, with two oblique, lateral and marginal keels.
Among all the species belonging to Pleurothallis subsection Macrophyllae-Fasciculatae, P. carpishensis is most similar morphologically to P. radula from Costa Rica ( Figure 7 View FIGURE 7 ), sharing characteristics such as the ovate to lanceolate leaves, deeply cordate at the base with the basal lobes sometimes overlapping, the solitary flowers, erect dorsal sepal, reflexed, single-veined petals and the thick and fleshy lip. However, P. carpishensis is easily differentiated from P. radula by the smaller plants, up to 23 cm tall (vs. up to 50 cm tall), ovary 2.7–3.0 mm long (vs. 4.0–6.0 mm long), dorsal sepal 8.0– 10.3 mm long (vs. 12–14 mm long), synsepal 8.0–10 × 4.5–4.8 mm (vs. 12.0–14.0 × 7.0 mm), petals 4.5–6.0 mm long (vs. 8.0– 9.5 mm long), lip with a callus widely ovate, emarginate, papillose-verrucose, 1.7 × 2.3 mm (vs. glabrous, callus 1 mm long), without glenion (vs. glenion slightly raised above callus) and the ciliate periphery of the stigmatic surfaces (vs. slightly papillate).
As in many species of Pleurothallidinae , the leaves of P. carpishensis show vegetative variability. Therefore, some individuals have leaves ranging from ovate to ovate-lanceolate, and occasionally falcate near the apex, with the apex acute to shortly acuminate, with the basal lobes cordate and sometimes overlapping ( Figure 8 A – B View FIGURE 8 ). However, a peculiar characteristic in P. carpishensis are the apices of the ramicaul, since these are curved, forming an angle of approximately 90°, before the leaf abscission layer ( Figure 8 C View FIGURE 8 ), unlike other Pleurothallis species of the same subsection, which generally have leaves attached to a non-angulate ramicaul.
Interestingly, in some species of subsection Macrophyllae-Fasciculatae, the single-flowered inflorescences are borne from an erect spathaceous bract ( Pupulin et al. 2021) [e.g. P. longipetala Bogarín & Belfort (2021: 265) , P. phyllocardia Reichenbach (1866: 97) , P. radula , P. rectipetala Ames & C.Schweinf. (1925: 32) ], that emerge from a horizontal to erect leaf or leaf deflexed toward the ramicaul. Nevertheless, in P. carpishensis the flower is borne from a depressed spathaceous bract that lies over a navicular concavity on the leaf ( Figure 8 View FIGURE 8 , red arrow). This concavity is as long as the flower which overlies it ( Figure 8 D View FIGURE 8 ). This makes the species unmistakable, compared to the other Macrophyllae-Fasciculatae occurring in Peru.
Since P. radula is not found in Peru, field botanists in that country need to distinguish P. carpishensis from the most morphologically similar species to be encountered in that country, in this case P. crucifera . Pleurothallis carpishensis is similar to P. crucifera in the coriaceous, ovate, acute leaves, the yellow-brown, glabrous flowers, that rest on the leaf and the long, acute petals ( Figure 9 View FIGURE 9 ). However, P. carpishensis differs from P. crucifera by the length of the ramicauls (8.0– 20.6 cm long vs. 5.0–10.0 cm long), arrangement and direction of the leaves (reflexed vs. spreading), with attenuate, tridentate (vs. acute) apex, the floral bracts long-conical in shape, 3.5–5.0 mm long (vs. tubular, 5.0–6.0 mm long), the pedicels 4.0–5.0 mm long (vs. 6.0–8.0 mm long), the flowers hyaline, yellow concolor (vs. light yellowish or brown flowers), arrangement and direction of the petals (porrect vs. spreading), the lip without glenion (vs. large glenion), and the stigma ventral, reniform (vs. apical, sub-rectangular), with margins of stigmatic surface ciliate margins (vs. papillose-erose margins).
Within Pleurothallis subsection Macrophyllae-Fasciculatae, most species exhibit a glenion at the base of the lip, located below the apically located viscidium of the pollinarium. Morphologically, the glenion appears to be a small secretory structure ( Wilson et al. 2016, Wilson et al. 2022) and we hypothesize that the glenion aids in pollinator positioning for optimal pollinarium acquisition and subsequent deposition on another flower. Yet surprisingly some species, such as P. carpishensis , P. minutilabia , P. rhopalocarpa , and the P. crateriformis complex appear to lack such structure. In P. rhopalocarpa , a very widely distributed species, the lateral margins produce copious quantities of liquid ( Figure 10 A – C View FIGURE 10 ) that is presumably a nectar reward, and this species may have evolved to permit more flexible pollinator positioning during pollinarium acquisition than occurs in species with a glenion. On the other hand, P. carpishensis neither possesses a glenion nor secretes nectar-like liquid from the medial and apical regions of the lip ( Figure 10 D View FIGURE 10 ), suggesting a unique pollination strategy yet to be elucidated. In addition, P. carpishensis exhibits ciliatepapillate margins around the stigmatic surfaces of the column ( Figure 3 E View FIGURE 3 ) and the petals ( Figure 3 C View FIGURE 3 ), both of which are rare in Macrophyllae-Fasciculatae. The ciliate papillae could be involved in secretion of volatiles involved in pollinator attraction, but determination of their function will require additional study.







| USM |
USM |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |