
Pareisactus evrostos, Párraga & Prieto-Márquez, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4555.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:4B7BCCAE-3C16-4D64-A19D-34C910279BF |
|
DOI |
https://doi.org/10.5281/zenodo.5936048 |
|
persistent identifier |
https://treatment.plazi.org/id/03C75868-3815-FFEF-FF23-FDE59290AAD0 |
|
treatment provided by |
Plazi |
|
scientific name |
Pareisactus evrostos |
| status |
sp. nov. |
Pareisactus evrostos sp. nov.
( Figs. 2 View FIGURE 2 and 3 View FIGURE 3 )
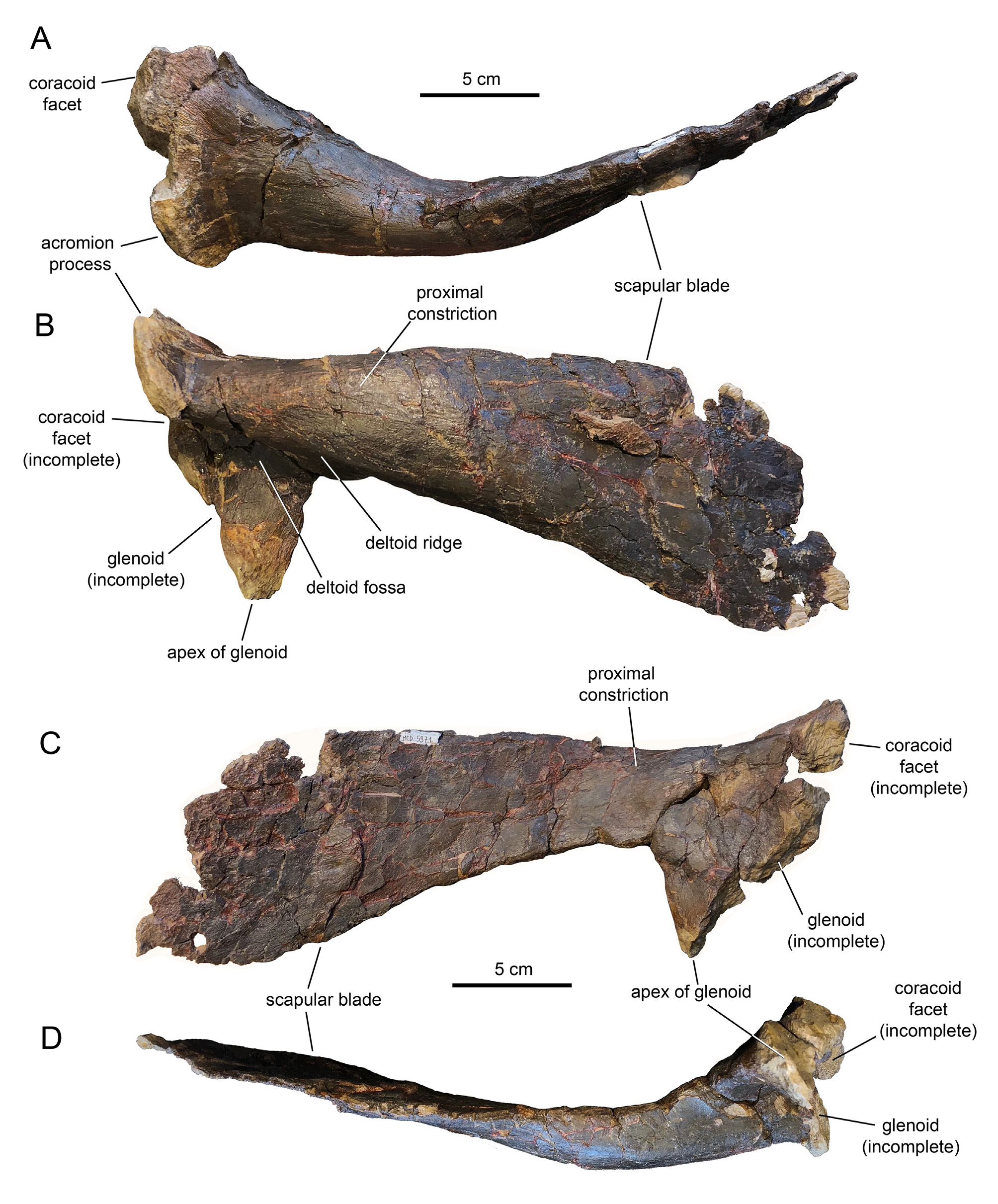
Holotype: MCD 5371 View Materials , a nearly complete left scapula.
Etymology: Eύρωστoς ( evrostos ) means ‘robust’ in Greek, in reference to the great mediolateral thickness of the proximal extent of the scapula and massive build of the deltoid ridge.
Type locality: Basturs Poble, near the village of the same name, Lleida Province, northeastern Spain ( Fig. 1 View FIGURE 1 ).
Type horizon: Upper lower Maastrichtian strata of the Conques Formation, Tremp Group, eastern Tremp Syncline, northeastern Spain ( Fondevilla et al. 2016) ( Fig. 1 View FIGURE 1 ).
Diagnosis. Basal iguanodontian ornithopod possessing the following autapomorphies: scapula with dorsoventral extent of proximal constriction (scapular ‘neck’) as deep as it is thick; lateral surface of proximal scapula under acromion process forming deep and thick roof over extremely deep deltoid fossa; long glenoid apex accounting for half of maximum depth of proximal region of scapula. Pareisactus evrostos differs from dryomorphan ornithopods in combining realtively long craniodorsally projected acromion process, craniocaudally short scapular ‘neck’, and glenoid apex that is strongly offset caudally from level of craniodorsal tip of the acromion process. It differs from other pre-dryomorphan ornithopods in the autapomorphies listed above.
Description. MCD 5371 is a nearly complete left scapula, missing part of the coracoid facet, the cranial margin of the glenoid, and a substantial part of the caudodorsal extent and caudoventral margin of the distal region of the scapular blade ( Fig. 2 View FIGURE 2 ; Table 1). At about two-fifths of the scapular length, there is a fracture that crosses the bone dorsoventrally and that has exaggerated the longitudinal medial curvature of the scapula. The lateral surface of the scapula is strongly convex dorsoventrally, particularly along its thickest proximal extent ( Fig. 2B View FIGURE 2 ), becoming flatter distally along the scapular blade. In contrast, the medial surface of the scapula is flat throughout the entire length of the element ( Fig. 2C View FIGURE 2 ).
The dorsal region of the proximal region of the scapula of Pareisactus evrostos is greatly expanded mediolaterally, so that the proximal constriction (‘scapular neck’) is as dorsoventrally deep as it is mediolaterally thick ( Fig. 2A and B View FIGURE 2 ). The deltoid fossa is extremely deep and its lateral surface projects ventrally to form the apex of the glenoid ( Fig. 2B View FIGURE 2 ). This lateral surface under the deltoid ridge leading into the glenoid apex is remarkably long, accounting for half of the maximum depth of the proximal region of the scapula including the acromion process ( Fig. 2B View FIGURE 2 ). The deltoid fossa is roofed by a prominent deltoid ridge. This ridge extends caudoventrally becoming continuous with the ventral margin of the scapular blade ( Fig. 2B View FIGURE 2 ). At the cranial end of the scapula, the dorsal margin curves dorsally and laterally to form a mediolaterally broad acromion process ( Fig. 2A and B View FIGURE 2 ). This process projects craniodorsally, as well as craniolaterally, and its lateral profile appears nearly parallel to the eroded cranial margin of the glenoid ( Fig. 2B View FIGURE 2 ). The acromion process of P. evrostos extends above the dorsal margin of the scapula a shorter distance than in other ornithopods such as Orodromeus makelai ( Scheetz 1999) , Oryctodromeus cubicularis ( Varricchio et al. 2007) , Haya griva ( Makovicky et al. 2011) or Camptosaurus dispar ( Gilmore 1909), but a longer distance than in Hadrosauriformes ( Brett-Surman & Wagner 2007) and at least some basal styracosternans like Bolong yixianensis ( Wu 2010) ( Fig. 3 View FIGURE 3 ). As in non-hadrosauriform iguanodontians (e.g., McDonald 2011; Norman 2011), and particularly more distant outgroups such as rhabdodontids (e.g., Chanthasit 2010) or jeholosaurids (e.g., Butler et al. 2011), the apex of the acromion process is strongly offset cranially relative to the position of the apex of the glenoid ( Fig. 3 View FIGURE 3 ).
Caudally, the paddle-shaped scapular blade gradually expands dorsoventrally, more ventrally than dorsally, while progressively thinning mediolaterally to become a delicate lamina at its distal end. As preserved, the maximum distal expansion of the scapular blade is 90 percent of the maximum depth of the proximal end (from the apex of the acromion process to the apex of the glenoid) and two and half times deeper than the proximal constriction. The total length of the scapula (from the apex of the acromion process to the distal end of the scapular blade) is only 2.4 times the maximum depth of the proximal end. Such relatively short scapulae, with length/ proximal depth ratios less than three, are typically found in styracosternan-outgroup ornithopods such as Orodromeus makelai ( Scheetz 1999) , jeholosaurids ( Butler et al. 2011; Makovicky et al. 2011), rhabdodontids ( Chanthasit 2010) or Camptosaurus dispar ( Gilmore 1909).
Relationships of Pareisactus evrostos . The phylogenetic position of Pareisactus evrostos was inferred using Parsimony. The taxonomic sample included 52 ornithopod species and Lesothosaurus diagnosticus as outgroup taxon. We included the unnamed lambeosaurine ( Fondevilla et al. 2018) from the Basturs Poble bonebed to the analysis. The subadult elements of the Basturs Poble lambeosaurine were scored only for those characters the states of which do not vary during ontogeny, according to Prieto-Márquez (2011) and Prieto-Márquez & Guenther (2018). The data set (see Supporting Material) consisted of the 134 morphological characters of Madzia et al. (2018), to which six more characters were added (totaling 94 cranial and 46 postcranial discrete morphological characters). Multistate characters containing states that are not mutually exclusive, following a natural morphocline, were ordered. This criterion allows for ‘crediting’ shared intermediate states. The search for the optimal tree(s) was conducted in TNT version 1.1 ( Goloboff et al. 2008). A heuristic search of 10,000 replicates using random additional sequences was performed, followed by branch swapping by tree-bisection-reconnection holding ten trees per replicate. Bremer support ( Bremer, 1988) was assessed by computing decay indices ( Donoghue et al. 1992) using TNT. Bootstrap proportions ( Felsenstein 1985) were also calculated using TNT, setting the analysis for 5,000 replicates using heuristic searches, in which each search was conducted using random additional sequences with branch-swapping by subtree pruning and regrafting and 25 replicates.
The phylogenetic analysis resulted in 12 most parsimonious trees of 504 steps each (C.I. = 0.45, R.I. = 0.80); the best score was found in 904 of the 10,000 replicates. Pareisactus evrostos was recovered as a basal iguanodontian, nested within Rhabdodontidae as the sister taxon to Rhabdodon priscus ( Fig. 3 View FIGURE 3 ). Pareisactus and Rhabdodon are unambiguously united by the presence of a well-demarcated deltoid ridge. However, this sister relationship is weakly supported, as indicated by the low decay indices and Boostrap proportions below 50%. We tested how many more steps would less parsimonious trees have when forcing Pareisactus to be placed in other areas of the iguanodontian tree. In particular, moving Pareisactus outside Iguanodontia caused the tree length to increase in one more step, as it did when positioned as sister to Thescelosaurus . Notably, tree length increased in seven, six and again six steps when placing Pareisactus within Hadrosauriformes, Hadrosauroidea and Hadrosauridae , respectively.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |