
Percina sipsi, Williams & Neely
|
publication ID |
https://doi.org/ 10.5281/zenodo.178071 |
|
publication LSID |
lsid:zoobank.org:pub:9DCBB385-6157-47E1-83C7-738E416D609E |
|
DOI |
https://doi.org/10.5281/zenodo.6237371 |
|
persistent identifier |
https://treatment.plazi.org/id/03C587A3-FFC5-0038-FF7F-FEED2DE5FEC7 |
|
treatment provided by |
Plazi |
|
scientific name |
Percina sipsi, Williams & Neely |
| status |
sp. nov. |
Percina sipsi, Williams & Neely View in CoL , new species
Bankhead Darter
( Fig. 2 View FIGURE 2 b)
Holotype. UF 165737, 46 mm SL, Sipsey Fork at Alabama Hwy 33, 3 km NE of Rock Springs, Winston County, Alabama (34°13’05”N; 87°22’09”W), 20 January 2003.
Paratypes. Black Warrior River drainage—Sipsey Fork system
Alabama: Lawrence County: TU 163064 (1; 43 mm) Borden Creek, tributary to Sipsey Fork, 29 April 1992. TU 168262 (1; 50 mm) Borden Creek at Forest Road, Sipsey Wilderness, Section 12, 12 September 1993. UAIC 6427.03 (1; 39 mm) Borden Creek at Forest Service Route 224, Bankhead National Forest (34°18'34"N; 87°23'42"W), 19 April 1981. Winston County: AUM 18786 (6; 35–45 mm) West Fork Sipsey Fork at Forest Service Road 234, 8.8 km NNE of Double Springs (34°13'32"N; 87°22'37"W), 3 June 1979. UF 103304 (9; 37–49 mm) same locality, 17 April 1995. TU 83137 (7; 16–46 mm) same locality, 3 July 1971. UAIC 4329.21 (5; 39–46 mm) West Fork Sipsey Fork, 21.6 km ENE of Haleyville (34°17'03'N; 87°23'56"W), 2 November 1978. INHS 48682 (7; 29–47 mm) Sipsey Fork at Alabama Hwy 33, 3 km NE of Rock Springs (34°13’05”N; 87°22’09”W), 13 August 1998. UAIC 10390.08 (6; 31–40 mm) same locality, 20 August 1986. UF 165738 (5; 38–48 mm) same locality, 20 January 2003. UMMZ 194316 (9; 36–40 mm) Sipsey River at Low Pressure Bridge, about 6.4 km E of Alabama Hwy 195, about 8.8 km NNE of Double Springs (T9S, R8W, S23), 15 October 1971. USNM 211256 (35; 32–44 mm) Sipsey River, about 4 km W of Grayson and about 16 km NNE of Double Springs, (T9S, R8W, S10), 29 October 1971.
Additional material (nontypes).
Black Warrior River drainage—Sipsey Fork system
Alabama: Lawrence County: AUM 27232 (1) Borden Creek at gravel road, Sipsey Wilderness, 22.2 air km SW of Moulton, 30 August 1991. Winston County: AUM 27043 (4) Sipsey Fork at Alabama Hwy 33 (34°13’05”N; 87°22’09”W), 27 November 1990. TU 83136 (1) Sipsey Fork, 8.5 km E of Ashridge, 1 km above Lake Lewis Smith (T9S, R8W, S33), 3 July 1971. UAIC 1695.21 (8) West Fork Sipsey Fork, 22.5 km ENE of Haleyville (34°16'24"N; 87°22'28"W), 12 July 1978. UAIC 4323.18 (1) West Fork Sipsey Fork, 20.6 km ENE of Haleyville (34°17'56"N; 87°24'55"W), 2 November 1978. UAIC 10274.02 (3; 39–48 mm) West Fork Sipsey Fork at Forest Service Road 234, 8.8 km NNE of Double Springs (34°13'32"N; 87°22'37"W), 2 March 1992.
Material used in molecular analysis.
Black Warrior River drainage
Alabama: Winston County: STL 1406.01 (4) Sipsey Fork at Alabama Hwy 33, 3 km NE of Rock Creek (34°13'05"N; 87°22'09"W), 20 January 2003, EF613207 View Materials , EF613208 View Materials , EF613209 View Materials , EF613210 View Materials
Diagnosis. Percina sipsi is distinguished from all other described species of Percina by a combination of the following characteristics: absence of bright colors on body and fins of adults; no orange band in spinous dorsal fin; no broad vertical bands on body extending dorsally across the back joining those of the other side; 7–11 lateral blotches fused into continuous dark brown to black lateral stripe with undulating margins; lateral stripe continuous with large, somewhat quadrate basicaudal blotch that extends onto base of caudal fin rays; small, dark blotch on upper and lower portion of caudal fin base, dorsal blotch typically darker; body below lateral stripe white to cream colored, without dark blotches, becoming dusky in breeding males; suborbital bar absent or very poorly developed; lateral line complete, typically no pored scales on base of caudal fin; males with row of modified scales on midline of belly and one or two modified scales between base of pelvic fins; modified breast scale absent; nuptial tubercles absent; anal fin of breeding males not excessively elongate; males without caudal keel as a ventral extension of the caudal peduncle; snout does not project beyond anterior margin of upper jaw; broad premaxillary frenum present; serrae on margin of preopercle absent; branchiostegal membranes very narrowly joined to overlapping.
Percina sipsi is distinguished from the other two species described herein by a combination of the following characters: dorsal saddles usually present but may be poorly developed, typically consisting of quadrate blotches on the posterior portion of nape, under posterior end of spinous dorsal fin, and the soft dorsal fin; dorsum with dusky brown reticulations above lateral stripe; cheeks and opercles scaled; nape typically naked, occasionally partially scaled; breast naked, occasionally with a few embedded cycloid scales.
Description. Percina sipsi is one of the two smallest species in the genus Percina , rarely exceeding 50 mm SL ( P. brevicauda , maximum SL is 50 mm, see Suttkus et al. 1994). Shape of the head and body are illustrated in Figure 2 View FIGURE 2 b. Frequency distribution of scale, fin ray and vertebral counts given in Tables 1–8 View TABLE 1 View TABLE 2 View TABLE 3 View TABLE 4 View TABLE 5 View TABLE 6 View TABLE 7 View TABLE 8 . Degree of nape squamation presented in Table 9 View TABLE 9 and posterior extent of development of pored lateral line scales given in Table 10 View TABLE 10 . Proportional measurements presented in Table 11 View TABLE 11 . Body terete, moderately elongate, snout moderately long, about equal to orbit in length, with a well-developed frenum. Preopercular margin entire and branchiostegal membranes separate or very narrowly joined. Total lateral line scales 56–72, usually 60–69; transverse scale rows 14–19, usually 16 or 17; caudal peduncle scales 18–22, usually 19–21; dorsal spines 12– 15, usually 13 or 14; dorsal soft rays 9–11, usually 9 or 10; anal soft rays 6–8, usually 8; pectoral rays 13–14, usually 13; vertebrae 40–42, usually 41.
Lateral line complete but occasionally an individual will have 1–3, usually one or two, unpored scales before the posterior edge of the hypural plate or a pored scale on the base of the caudal fin ( Table 10 View TABLE 10 ). Embedded scales present on the upper three quarters of the cheek and opercle. Nape usually naked, occasionally with some embedded scales ( Table 9 View TABLE 9 ). Breast and prepectoral area naked, rarely individuals with a few embedded scales. Belly typically scaled with the exception of the area immediately posterior of pelvic fin base. A row of enlarged, modified ctenoid scales present on midline of belly but ends a few scale rows posterior to pelvic fin base. Typically one or two modified scales present between the bases of pelvic fins. The large modified breast scale at the anterior margin of the pelvic girdle of some Percina is absent in P. s i p s i. In females the scales along the midline of the belly and laterally one to two rows are often embedded. Midventral row of modified scales on the belly of females is greatly reduced in development and number compared to males and is absent in some individuals. There are usually one or two modified scales present between the bases of the pelvic fins. Breeding tubercles and the thickened ridges along margin of anal fin rays of some males are absent in females.
The general pattern of pigmentation in life is similar in males and females but females usually lack the dusky ground color present in males, most pronounced in breeding males. Sides of the body with 7–10 round to slightly oval, dark brown blotches which are typically connected by a lighter brown lateral stripe two or three scale rows wide. Some individuals appear to have smaller blotches between larger blotches but are obscured by the underlying lateral stripe. Lateral blotches are usually lighter in color and less connected in small individuals and females. Anteriorly the lateral stripe merges with the postorbital and preorbital bars. The lateral stripe terminates on the base of the caudal fin in a quadrate basicaudal blotch. There are also dark blotches on the caudal fin base above and below the basicaudal blotch. These blotches vary in size and intensity but the upper blotch is usually more distinct than the lower. Above the lateral stripe the dorsum is variously marked with tan to light brown reticulations and blotches, varying in size and intensity. Most reticulations are formed by darkly pigmented scales and are 1–3 scale rows wide. Some individuals have reticulations that are aligned in a pattern two or three scale rows above the lateral stripe while others have no discernible pattern. Along the midline of the dorsum there are up to nine tan to light brown poorly developed saddles and when present typically consist of quadrate blotches. These blotches are often irregular in shape and vary in their placement and development. Their alignment usually consists of one blotch on the nape just anterior to the dorsal fin origin and if a second is present it is located anteriorly on the nape, three blotches under the spinous dorsal, one near the junction of the spinous and soft dorsal fin, two under the soft dorsal fin, and two on the caudal peduncle. Below the lateral stripe the body is usually devoid of blotches, spots, or reticulations but may be dusky on breeding males. On the head, the cheeks and opercles below the postorbital stripe lack dark pigment but may be dusky on breeding males. A well-developed suborbital bar is absent but a cluster of large melanophores may be present on the midventral margin of the orbit. The breast is without any dark pigment but there is typically a cluster of melanophores present on the anterior prepectoral region.
The spinous dorsal fin of males has a thin, dusky margin, a clear submarginal band, and a dark basal band which is usually darker posteriorly. The soft dorsal, anal, and caudal fins are usually clear but may be dusky in breeding males, with a slight intensification of pigment towards the margin. The pelvic and pectoral fins are usually clear but are dusky in breeding males.
Distribution. Percina sipsi is known only from the Sipsey Fork of the Black Warrior River in the Bankhead National Forest in northwestern Alabama ( Fig. 3 View FIGURE 3 ). It occurs in Borden Creek in Lawrence County, and Brushy Creek, Caney Creek, and Sipsey Fork (recorded as West Fork Sipsey River on some maps) in Winston County. While the current known range of P. sipsi is very limited and confined downstream by Lewis Smith Reservoir, historically it is possible that the species ranged farther downstream in the Sipsey Fork and conceivably in the Locust and Mulberry forks of the Black Warrior River, which are all located in the Cumberland Plateau physiographic province ( Fig. 3 View FIGURE 3 ). Riverine habitat in the lower reaches of the Sipsey Fork was destroyed in 1960 by the Alabama Power Company impoundment behind Lewis Smith Dam. The current distribution of P. sipsi represents the most restricted range of any known species of Percina .
Ecology. Percina sipsi inhabits creeks and small rivers ranging in size from 5–40 m in width and 0.25 to 2 m in depth. The species occurs in clear water over sand and fine gravel, usually in association with leaf packs and/or woody debris, but occasionally over the broad expanses of open bedrock which are abundant in parts of the mainstem Sipsey Fork.
Conservation status. Ramsey (1976) published the first conservation status review of Percina sipsi and reported it to be a threatened species. Although the assigned conservation status category has varied in subsequent evaluations ( Deacon et al. 1979; Ramsey 1984; Williams et al. 1989; Warren et al. 2000), all authors considered the Bankhead Darter’s continued existence to be in a precarious situation. In the most recent assessment of conservation status of Alabama wildlife, Kuhajda (2004) reported it as a species of highest conservation concern. The entire range of this species is in the Bankhead National Forest, providing some level of protection. We consider P. s i p s i to be highly endangered based on its restricted distribution, rarity within the occupied range, habitat vulnerability, and absence of downstream habitat for future recovery. Percina sipsi is extremely vulnerable and needs continuous monitoring and proactive management actions to prevent extinction.
Etymology. The specific name, sipsi , is the Chickasaw-Choctaw Indian name for poplar or cottonwood tree, and is the origin of “Sipsey” in the stream name, Sipsey Fork of the Black Warrior River ( Foscue 1989), to which this species is endemic. The common name, Bankhead Darter, is in reference to the William B. Bankhead National Forest which encompasses the range of the species.
Comparison with congeners. In the Sipsey Fork, a tributary to the Black Warrior River, Percina sipsi occurs with five species of Percina : P. kathae , P. maculata , P. nigrofasciata , P. sciera and P. shumardi . Prior to construction of the impoundments on the Black Warrior River it may have occurred sympatrically with a sixth species, P. brevicauda . Percina sipsi is distinguished from P. kathae which has the snout projecting beyond the anterior margin of the upper jaw, numerous narrow vertical bars on the dorsum and a well-developed basicaudal spot. It differs from P. maculata which has a well-developed subocular bar and basicaudal spot and a dark blotch on the base of the anterior 3–4 membranes of the spinous dorsal fin. Males of P. maculata typically have a well-developed modified breast scale. Percina sipsi can be distinguished from P. nigrofasciata which has vertically elongate lateral blotches and moderately joined gill membranes. Percina sipsi differs from P. shumardi in having a well-developed frenum and a row of modified scales along the midline of the belly of males. Breeding males of P. sipsi also lack the elongation of the anal fin and breeding tubercles on the anal, caudal, and pelvic fins that are characteristic of P. shumardi . Percina sciera is superficially very similar to P. sipsi and the two are difficult to distinguish in the field. The most reliable characters to distinguish the two species are the more moderately joined gill membranes and typically serrate preopercle margin in P. sciera compared to the narrowly joined gill membranes and smooth preopercle margin of P. s i p s i. Nape squamation is also a helpful character to distinguish the two species, with P. s c i e r a having exposed and/or embedded scales over the entire nape while P. s i p s i is typically naked or only has a few embedded scales. Percina sipsi is a small species, 50 mm maximum known SL, while P. s c i e r a often exceeds 50 mm SL. The standard length of 58 mm for P. s i p s i (UAIC 10274.02) reported in Boschung and Mayden (2004) should read 48 mm. If P. sipsi and P. brevicauda were found to occur sympatrically they could be distinguished by the presence of a frenum in P. sipsi and a black basicaudal spot in P. brevicauda .
Holotype. UF 165735, 47 mm SL, Turkey Creek at unnumbered county road, about 2.9 km W of Burwell, Carroll County, Georgia (33°35’45”N; 85°13’36”W), 19 January 2003.
Paratypes. Tallapoosa River drainage
Alabama: Chambers County: AUM 23918 (7; 38–51 mm) Tallapoosa River, 2.4 km SSE of Wadley(33°06'07"N; 85°33'25"W), 1 November 1982. UAIC 8525.07 (4; 35–51 mm) same locality, 14 May 1986. UAIC 8526.09 (6; 43–53 mm) same locality, 14 May 1986. Clay County: TU 157782 (3; 44–48 mm) Enitachopco [Enitochope] Creek at Alabama Hwy 9, 4.5 km SSW of Ashland (33°14'24"N; 85°51'34"W), 12 April 1990. UMMZ 185373 (5; 45–54 mm) same locality, 21 September 1963. Cleburne County: UF 98484 (5; 38–47 mm) Snake Creek at County Route 13, about 5.6 km SW of the center of Heflin (33°35'49"N; 85°37'21"W), 16 July 1992. Elmore County: TU 157794 (6; 29–50 mm) Gold Branch, 2.9 km E of Alabama Hwy 63 (32°41'35"N; 86°00'26"W), 12 April 1990. Randolph County: AUM 23824 (19; 32–49 mm) Crooked Creek, 7.4 km NW of Malone (33°14'36"N; 85°38'36"W), 27 September 1982. NCSM 44986 (6; 45– 55 mm) Tallapoosa River, 6.4 km NNW of Wadley (33°10'27"N; 85°35'04"W), 26 March 1991. NCSM 44987 (7; 36–54 mm) same locality, 4 September 1996. UAIC 8508.14 (2; 38–53 mm) same locality, 4 September 1996. UAIC 10830.20 (5; 40–44 mm) same locality, 26 March 1991. USNM 218508 (6; 34–54 mm) Jones Creek at US Hwy 431, approximately 0.6 km N of Roanoke (33°10'32"N; 85°23'21"W), 17 April 1978. USNM 322736 (5; 43–53 mm) Cornhouse Creek at Alabama Hwy 431 (33°14'20"N; 85°26'31"W), 23 March 1970. UT 91.2143 (16; 30–50 mm) Little Tallapoosa River at US Hwy 431 about 6.4 km N of Wedowee (33°22'03"N; 85°28'36"W), 16 May 1981. Tallapoosa County: INHS 57809 (5; 40–51 mm) Josie Leg Creek at Alabama Hwy 22, 6.1 km SW of Perryville (33°00'07"N; 85°49'58"W), 12 March 1989. NCSM 44985 (4; 39–43 mm) Hillabee Creek, at Alabama Hwy 22, about 9.7 km NE of Alexander City (32°59'07"N; 85°51'41"W), 22 March 1995. UF 91625 (14; 35–53 mm) same locality, 6 April 1992. TU 168037 (4; 44–52 mm) Emuckfaw Creek, 4.8 km W of Daviston (33°03'18"N; 85°41'41"W), 10 July 1993. TU 168051 (1; 51 mm) same locality, 10 July 1993. UAIC 6787.14 (3; 38–54 mm) Emuckfaw Creek 5.3 km SSE of New Site (32°59'43"N; 85°44'58"W), 3 June 1985. UAIC 8518.09 (5; 40–51 mm) same locality, 11 March 1985. UAIC 8476.20 (16; 37–51 mm) Tallapoosa River, 10.8 km SSW of Daviston (32°57'52"N; 85°41'52"W), 22 October 1986. UAIC 8510.10 (3; 35–51 mm) Tallapoosa River, 5.6 km SSE of New Site (32°59'06"N; 85°45'09"W), 11 March 1985. UF 165730 (3; 44–49 mm) Wind Creek at second bridge on Alabama Hwy 50 upstream of mouth (32°40’42”N 85°52’34”W), 18 January 2003. UF 165733 (5; 35–40 mm) Emuckfaw Creek at Alabama Hwy 49, just upstream of Horseshoe Bend NHP (32°59’40”N 85°44’59”W), 18 January 2003. USNM 199818 (14; 35–48 mm) Unnamed tributary of Sougahatchee Creek at Alabama Hwy 49, 19 km N of Tallassee (32°38'04"N; 85°47'48"W), 10 April 1965.
Georgia: Carroll County: UF 165736 (6; 43–48 mm) Turkey Creek at unnumbered county road, about 2.9 km W of Burwell (33°35’45”N 85°13’36”W), 19 January 2003. Haralson County: GMNH 1548 (3; 45– 57 mm) Tallapoosa River at County Road 130, 23 May 1981. UF 15856 (5; 35–53 mm) North Fork of Walkers Creek at Georgia Hwy 120, 6.1 km SW of Buchanan, 23 April 1968. UF 165731 (8; 43–51 mm) Beech Creek at Georgia Hwy 120, 5.2 km SW of Buchanan (33°45’46”N 85°13’22”W), 19 January 2003. Paulding County: UF 165732 (3; 51–58 mm) McClendon Creek at Georgia Hwy 101, 5 km SSE of Beulah (33°51’45”N 84°59’35”W), 19 January 2003. USNM 218470 (10; 38–50 mm) same locality, 17 April 1978.
Additional material (nontype).
Tallapoosa River drainage
Alabama: Chambers County: AUM 22087 (4) Tallapoosa River, 2.7 km SSE of Wadley (33°06'05"N; 85°33'23"W), 8 December 1981. Clay County: AUM 15034 (7) White Oak Creek, 1.9 km S of Cragford (33°14'05"N; 85°40'20"W), 26 July 1977. UAIC 2153 (7) Ketchepedrakee Creek, 0.5 km S of Dempsey (33°26'29"N; 85°46'15"W), 6 July 1966. UF 15444 (5) Little Hillabee Creek, 17.7 km NE of Goodwater, 12 September 1963. UF 93917 (1) Creek formed by confluence of Horse Creek and Cave Creek, 2 km SE of Union (33°26'01"N; 85°47'12"W), 16 July 1992. Cleburne County: UAIC 6625.08 (3) Cahulga Creek at Alabama Hwy 9 (33°36'19"N; 85°36'09"W), 11 April 1982. Lee County: AUM 8677 (2) Sougahatchee Creek, about 3.4 km NW of Loachapoka (32°37'10"N; 85°38'01"W), 17 October 1950. Randolph County: AUM 23298 (6) Cornhouse Creek, 4.2 km NE of Malone (33°13'26"N; 85°32'54"W), 8 June 1982. UAIC 6793.22 (9) same locality, 21 July 1983. AUM 23794 (22) Crooked Creek, 7.4 km NW of Malone (33°14'36"N; 85°38'36"W), 18 August 1982. AUM 23962 (29) same locality, 8 November 1982. UAIC 8483.10 (12) same locality, 20 February 1985. UAIC 8487.15 (20) same locality, 7 November 1985. UAIC 8496.12 (5) same locality, 3 April 1986. UAIC 8512.12 (20) same locality, 21 January 1986. UF 99383 (2) Hurricane Creek at County Route 26, about 9.6 km NNW of Wadley (33°10'31"N; 85°35'54"W), 25 May 1995. UF 104096 (1) same locality, 2 March 1996. Tallapoosa County: JDW 95-15 (4) Hillabee Creek at Alabama Hwy 22, 10 km NE of Alexander City (32°59'07"N; 85°51'41"W), 22 March 1995. UAIC 6418.11 (5) same locality, 16 November 1980. UAIC 8494.09 (4) Tallapoosa River, 6 km SSE of New Site (32°59'06"N; 85°45"09"W), 11 March 1985. UAIC 8522.12 (6) same locality, 8 May 1986. UAIC 8495.21 (3) Emuckfaw Creek, 5.3 km SSE of New Site (32°59'43"N; 85°44'58"W), 20 February 1986.
Georgia: Paulding County: UF 165650 (1) Thomasson Creek at Georgia Route 136, 3 km WSW of Yorkville (33°55'01"N; 85°01'44"W), 1 May 1990.
Materials used in molecular analysis.
Tallapoosa River drainage
Alabama: Tallapoosa County: STL 731.02 (1) Wind Creek at second bridge on Alabama Hwy 50 upstream of mouth, below old mill dam (32°40'42"N; 85°52'34"W), 18 January 2003, EF613213 View Materials . STL 732.01 (1) Emuckfaw Creek at Alabama Hwy 19 just NW of Horseshoe Bend (32°59'40"N; 85°44'59"W), 18 January 2003, EF613214 View Materials .
Georgia: Carroll County: STL 1383.01 (1) Turkey Creek at unnumbered county road 3 km NW of Burwell (33°35'45"N; 85°13'36"W), 19 January 2003, EF613211 View Materials . Haralson County: STL 1384.01 (1) Beech Creek at Georgia Hwy 120, 5.2 km SW of Buchanan (33°45'46"N; 85°13'22"W), 19 January 2003, EF613215 View Materials . Paulding County: STL 1385.01 (1) McClendon Creek at Georgia Hwy 101, 5 km SSE of Beulah (33°51'45"N; 84°59'35"W), 19 January 2003, EF613212 View Materials .
Diagnosis. Percina smithvanizi is distinguished from all other described species of Percina by a combination of the following characteristics: absence of bright colors on body and fins of adults; no orange band in spinous dorsal fin; no broad vertical bands on body extending dorsally across the back joining those of the other side; 7–11 lateral blotches connected to form a continuous dark brown to black lateral stripe with undulating margins; lateral stripe continuous with large, somewhat quadrate basicaudal blotch, which extends onto base of caudal fin rays; a small dark blotch on upper and lower portion of caudal fin base, dorsal blotch typically darker; suborbital bar absent or very poorly developed; lateral line complete, typically no pored scales on base of caudal fin; males with row of modified scales on midline of belly and one or two modified scale between base of pelvic fins; modified breast scale absent; nuptial tubercles absent; anal fin of breeding males not excessively elongate; males without caudal keel as a ventral extension of the caudal peduncle; snout does not extend beyond anterior margin of upper jaw; broad premaxillary frenum present; serrae on margin of preopercle absent; branchiostegal membranes very narrowly joined to overlapping.
Percina smithvanizi is distinguished from the other two species described herein by a combination of the following characters: dorsal saddles usually present, typically consist of quadrate blotches on the posterior portion of nape, under posterior end of spinous dorsal fin, and anterior portion of soft dorsal fin; dorsum with dusky brown reticulations above lateral stripe; scales present on nape, cheeks, opercles and breast. Body depth variable among all three species, but average body depth relative to SL is deeper in P. smithvanizi than congeners described herein (see Morphometric comparisons).
Description. A moderately small species of Percina , the largest specimen examined is a male 63 mm SL. Frequency distribution of scale, fin ray and vertebral counts given in Tables 1–8 View TABLE 1 View TABLE 2 View TABLE 3 View TABLE 4 View TABLE 5 View TABLE 6 View TABLE 7 View TABLE 8 . Lateral line usually complete, but often has an unpored scale before the posterior edge of the hypural plate. Less frequently, a single pored lateral line scale present on the caudal fin base. Anteriormost portion of belly often naked, but midventral row of belly scales usually present and composed of modified ctenoid scales in both sexes, but usually more enlarged in males. Prepectoral region invested with exposed scales, as are the opercles and upper threefourths of the cheeks. Nape fully covered with scales, usually embedded anteriorly, but often exposed ( Table 9 View TABLE 9 ). Posterior portion of breast with embedded or exposed scales, but naked anteriorly. Enlarged ctenoid scales present between pelvic fin bases, but usually absent on breast on the anterior margin of the pelvic bones. Total lateral line scales 57–71, usually 60–68; transverse scale rows 12–18, usually 14–16; caudal peduncle scales 17–23, usually 19–21; dorsal spines 11–14, usually 12 or 13; dorsal soft rays 9–11, usually 9 or10; anal soft rays 7–9, usually 8; pectoral rays 11–14, usually 13; vertebrae 40–42, usually 41.
The most prominent feature of pigmentation in adults is the blue-black, undulating lateral stripe, formed by the coalescence of 8–11 elongate-oval lateral blotches, and contrasting sharply with the light areas above and below. The lateral blotches are usually brownish and more separated in juveniles. A large, well-defined quadrate basicaudal spot is present as a continuation or slightly interrupted continuation of the lateral band, and extends onto the base of the central caudal fin rays. Smaller, usually less obvious, blotches are present at the bases of the upper and lower principal caudal rays, although the lowermost blotch is frequently absent or obsolescent. The dorsum above the lateral stripe is tan to yellowish, and is variously marked with alternating brownish saddles and intermediate blotches, which are rarely continuous across both sides of the dorsum.
There is usually a dark brown saddle present on the nape. Males possess dusky marginal and basal bands in the spinous dorsal fin but the pigment in the basal band is darker and more concentrated posteriorly. The caudal fin is vaguely marked with three or four light bars. Other fins in the male are dusky, without pronounced banding. The breast and belly are dusky. A cluster of melanophores is usually present on the anterior base of the pectoral fin. Preorbital and postorbital bars are well-developed, and appear continuous with the lateral stripe. The suborbital bar is usually absent, or is very faintly developed in males as a scattering of melanophores and extending directly downward and slightly forward from the ventral margin of the orbit. The upper lip is pigmented above to about half its length posteriorly. The lower surface of the head is usually white to cream color.
Females are similar in color to males, but lack the dusky ground color on the body and fins and dark bands in the spinous dorsal. The cheek, breast, and belly are white to cream colored, often with a scattering of darkedged scales present around the anal fin. The dorsal fin rays are irregularly marked with faint blotches. The pelvic, pectoral, and anal fins are clear to dusky.
Spawning coloration of males and females collected in early June was yellowish above the lateral stripe, which was blue-black in males but slightly less intense in females, and some yellowish color was present immediately below the lateral stripe. A very pale greenish sheen was present on the lower sides in females. The rays of the dorsal, pectoral, and caudal fins were pale yellowish, the pelvic and anal rays colorless. The top of the snout and head were variously mottled or sprinkled with dusky, brown, orange, or yellow color. A brassy patch was present on the anterior portion of the opercles in adults, diminished in size in juveniles, and all specimens possessed some light blue iridescent pigment on the lower jaw and ventral surface of the head.
Nuptial tubercles were absent, but males in spawning condition had ridged, white epidermal thickenings on the anal rays, which may function as reproductive contact structures.
Distribution. Percina smithvanizi is restricted to streams draining the region above the Fall Line in the Tallapoosa River system ( Fig. 3 View FIGURE 3 ). It is widely distributed above the Fall Line except it is absent in upper reaches of the Little Tallapoosa River in Georgia (see Conservation status).
Ecology. Percina smithvanizi is common in the Tallapoosa drainage in the Tallapoosa River proper (width 75–125 m, depth 0.5–1 m) as well as tributaries (width 4–5 m, depth 0.2–1 m and larger). Individuals occur in clear water flowing over sand, gravel, rubble, and bedrock, in pools below riffles as well as riffles with moderate current, occasionally from margins of large rocks in the areas of fast flowing water. Areas of swift current yielded the larger specimens, while the smaller individuals were found in habitats with more moderate flow. Species most frequently captured with P. smithvanizi were Cottus sp. Tallapoosa Sculpin (Neely et al. in press), Etheostoma tallapoosae , Hypentelium etowanum , and P. palmaris . Other species commonly captured with P. smithvanizi included Campostoma oligolepis , Phenacobius catostomus , Cyprinella gibbsi , Noturus funebris , and N. leptacanthus . A detailed account of the ecology and biology of P. smithvanizi was reported by Wieland & Ramsey (1987).
Conservation status. Percina smithvanizi is a relatively common darter in undisturbed streams above the Fall Line in the Tallapoosa River. However, it has disappeared in disturbed streams and impoundments associated with mainstream dams on the Tallapoosa River. It has also disappeared from most of the headwaters of the Little Tallapoosa River in Georgia. The species was regarded as vulnerable by Freeman et al. (2005).
The upper two-thirds of the Tallapoosa River drainage was historically isolated from the lower one-third by a series of falls located at the Fall Line, near the present day town of Tallassee, Alabama. In the early 1900’s three dams were constructed on the Tallapoosa River in the vicinity of the Fall Line further isolating the upper and lower portions of the river system. The falls appear to have represented a barrier to upstream dispersal for many fishes, including Percina nigrofasciata . In a survey of fishes in the Tallapoosa River system ( Williams 1965) P. nigrofasciata was absent from most of the upper Tallapoosa River with the exception of the upper reaches of the Little Tallapoosa River in Georgia. Based on comparison of meristic and pigmentation characters with data from Crawford (1965), it appears that this population of P. nigrofasciata was introduced from the adjacent Chattahoochee River system where the species is common ( Williams 1965). Crawford (1965) reported a collection of P. nigrofasciata in the upper Tallapoosa River drainage, near the junction of the Tallapoosa and Little Tallapoosa rivers, but this record is based on specimens of P. smithvanizi . In the upper reaches of the Little Tallapoosa River system, the absence of P. smithvanizi in the presence of an introduced population of P. nigrofasciata suggests possible competitive displacement. If competitive displacement is occurring in the Little Tallapoosa River, the population of P. smithvanizi above the backwaters of R.L. Harris Reservoir may be at risk. Additional populations of P. nigrofasciata have been reported from tributaries of Lake Martin ( Boschung & Mayden 2004). These populations should be monitored to evaluate their movement and possible impact on P. smithvanizi and other darters.
P. kusha (n = 23) P. sipsi (n = 18) P. smithvanizi (n = 21) Etymology. We take great pleasure in naming this species for our good friend and colleague, Dr. William F. Smith-Vaniz, in recognition of his outstanding contributions to ichthyology in general and specifically for his authorship of the first book on the Freshwater Fishes of Alabama ( Smith-Vaniz 1968).
The common name, Muscadine Darter, is based on the dark purplish color of the lateral stripe in live individuals that is similar to the color of ripe Muscadine ( Vitis rotundifolia ) fruit in early fall. Muscadine vines occur along many of the more pristine streams inhabited by the Muscadine Darter.
% Variance 83.5 4.6 2.8 Cumulative % 87.9 90.6
Comparison with congeners. In the Tallapoosa River drainage above the Fall Line, Percina s mithvanizi occurs with three other species of Percina : P. kathae , P. nigrofasciata and P. palmaris . The pigmentation pattern of P. smithvanizi differs from P. kathae which has numerous narrow, vertical bars on the side and a welldeveloped basicaudal spot. Also the cone-shaped snout of P. kathae projects beyond the anterior margin of the upper jaw. Percina smithvanizi can be distinguished from P. nigrofasciata which has moderately joined gill membranes, vertically elongate lateral blotches and often has irregular shaped blotches on the belly. The broad saddles that extend across the back and bright colors of the fins and body of P. palmaris readily distinguish it from P. smithvanizi .
Morphometric comparisons
Proportional measurements for the three species of Percina are provided in Table 11 View TABLE 11 . Results of the sheared principal component analysis (PCA) are presented in Table 12 View TABLE 12 and Figures 4–5. In the PCA of all species, the first three components accounted for 91% of the morphometric variance. Variable loadings on the first principal component were positive and indicative of PC I as a general size factor ( Bookstein et al. 1985), and this component accounted for 83.5% of the total variance in the complete covariance matrix; values for PC II and PC III were 4.6% and 2.8%, respectively. The scatter of component scores on sheared PC II and sheared PC III revealed little differentiation among all three species (Fig. 4), with greatest overlap on both component axes between P. sipsi and P. smithvanizi . Heaviest loadings on sheared PC II were snout length and head length (both positive) and anal fin base length and body depth (both negative). Heaviest loadings on sheared PC III were body depth, pectoral fin length, pelvic fin origin to spinous dorsal fin origin (all positive), and anal and soft dorsal fin base lengths (both negative) ( Table 12 View TABLE 12 ).
To further compare shape differences between Percina kusha and P. smithvanizi , a second principal component analysis was done with the removal of measurements for P. sipsi from the data matrix. In this analysis, the first three components accounted for 88% of the morphometric variance, with loadings on PC I through PC III of 76.8%, 5.3%, and 2.6%, respectively. A scatter plot of component scores revealed moderate separation between the two species along PC II (Fig. 5). Variables that loaded most heavily on sheared PC II were anal fin base length and body depth (both positive) and snout length and head length (both negative); proportional differences between P. kusha and P. smithvanizi in these measurements were accompanied by substantial overlap in range values ( Table 11 View TABLE 11 ).
Based on results of the morphometric analysis, the combination of mensural data were of limited use in distinguishing specimens among the three Percina species. In spite of broad overlap for all morphometric characters among species, a slight difference in body depth between P. smithvanizi and the other two species was evident in a bivariate plot of log body depth versus log SL ( Fig. 6 View FIGURE 6 ). Analysis of covariance (ANCOVA; SPSS ver. 12.0) confirmed homogeneity of variance in the regression lines ( Table 13 View TABLE 13 ). Using the Bonferroni procedure to control for type I error across three comparisons (p <0.0167), post hoc contrasts indicated that P. smithvanizi differed significantly in adjusted mean log body depth from specimens of both P. kusha (F = 10.99; p = 0.002) and P. sipsi (F = 7.75; p = 0.007), but that the latter two did not differ from each other.
Our mtDNA data set consisted of 83 OTUs, including seven outgroup taxa. Thirty-three most-parsimonious topologies (L = 3972, CI = 0.22, RC = 0.13) were recovered in the unweighted MP analysis. The Bayesian topology ( Fig. 7 View FIGURE 7 ) was nearly identical to the consensus tree of the MP topologies, differing primarily in the degree of resolution of basal branches. Several features of these trees are worthy of note. While support for many terminal nodes was generally high, the basal relationships of the genus Percina are poorly resolved, consistent with the analyses of Near (2002) and Sloss et al. (2004). The subgenera Cottogaster , Imostoma , Percina and Swainia were all resolved as monophyletic groups, with strong support. Sufficient structure was resolved, however, to clearly support a sister-taxon relationship between P. kusha and P. smithvanizi , as well as a strongly supported clade consisting of P. aurolineata + (P. s c i e r a + P. s i p s i). The three taxa described herein were never recovered as a monophyletic group in any analyses. There was no consistent geographic structure observed in populations of either P. s i p s i or P. smithvanizi , however, populations of P. k u s h a from the Conasauga and Etowah River were always recovered as reciprocally monophyletic groups.
Populations of Percina sipsi , P. smithvanizi , and P. k u s h a displayed relatively low levels of variation ( Table 14 View TABLE 14 ). The five sampled specimens of P. k u s h a possessed three haplotypes; there were seven substitutions (four transitions and three transversions) and no indels. All sampled individuals from the Conasauga River shared a single haplotype, which differed from the two haplotypes sampled from the Etowah River by 0.53- 0.64% pairwise sequence divergence. The two Etowah haplotypes were 0.09% divergent from each other. The six sampled specimens of P. smithvanizi (representing the entire geographic range of the taxon) possessed four haplotypes; there were ten substitutions (five transitions and five transversions) and no indels. The haplotypes had pairwise sequence divergence values between <0.01% and 0.26%.
The four sampled specimens of Percina sipsi each had unique haplotypes; there were five substitutions (two transitions and three transversions) and no indels. Haplotypes had pairwise sequence divergence values between 0.18% and 0.35%. One specimen from the Sipsey Fork identified as P. s c i e r a on the basis of lateral blotching, fully scaled nape and a well-developed suborbital bar, had a haplotype that was very similar to that of P. s i p s i (and which only differed from P. s i p s i by between 0.26% and 0.44%). This specimen was resolved with P. s i p s i in all analyses.
Although P. s i p s i and P. s c i e r a are morphologically similar (see Comparison with congeners, above), recovery of a sister-taxon relationship between them was unexpected.
The specimens used in the molecular analysis were adult, morphologically "good" P. s i p s i and P. sciera . Our data do not allow us to determine whether this represents a natural relationship or an artifact of the mitochondrial locus examined. Considerable evidence exists for occasional mitochondrial introgression in darters ( Page et al. 2003, Mendelson & Simons 2006, N.J. Lang & J.M. Ray unpublished data), and we suspect that our recovery of P. s i p s i stems from introgression with P. sciera , but require additional nuclear data to test this hypothesis. Minimally, the haplotypic variation observed within P. s i p s i suggests that if the mitochondrial genome is introgressed with P. sciera , it is not the result of a recent event.

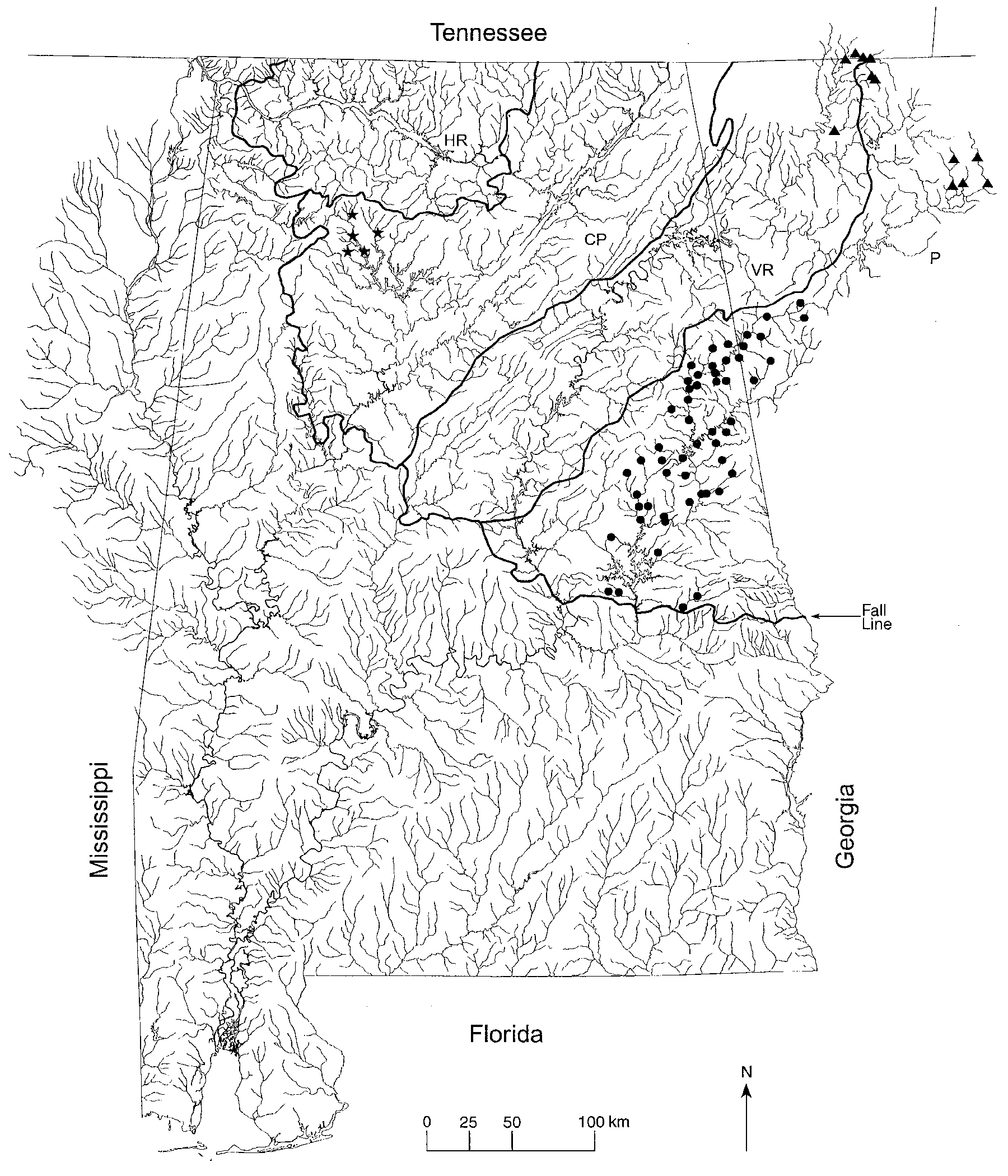


TABLE 10. Posterior extent of development of the pored lateral line scales in Percina kusha, P. sipsi and P. smithvanizi. Negative value indicates unpored scales before caudal fin base, positive value indicates pored scales on caudal fin base.
| Species | Posterior extent of pored lateral line scales | ||
|---|---|---|---|
| –3 –2 –1 0 +1 | N | ||
| P. kusha | |||
| Conasauga | – 2 7 14 2 | 23 | –0.5 |
| Etowah | – – 1 13 5 | 19 | 0.2 |
| P. sipsi | 1 4 17 33 2 | 57 | –0.5 |
| P. smithvanizi | 1 1 28 68 10 | 108 | –0.2 |
TABLE 11. Proportional measurements of Percina kusha, P. sipsi and P. smithvanizi, expressed in thousandths of standard length.
| Range | Mean | SD | Range | Mean | SD | Range | Mean | SD |
|---|---|---|---|---|---|---|---|---|
| standard length (mm) 43–65 | — | — | 37–50 | — | — | 43–57 | — | — |
| snout length 41–58 | 50 | 4.4 | 41–54 | 46 | 3.9 | 38–52 | 46 | 4.4 |
| eye diameter 47–73 | 64 | 6.3 | 56–70 | 63 | 3.3 | 57–71 | 65 | 3.9 |
| head length 219–258 | 235 | 9.2 | 206–243 | 229 | 10.0 | 212–257 | 231 | 11.7 |
| head depth 124–147 | 133 | 5.6 | 119–149 | 135 | 7.8 | 135–157 | 142 | 5.6 |
| occiput - pelvic fin origin 132–172 | 144 | 8.2 | 120–150 | 140 | 9.7 | 143–169 | 153 | 7.5 |
| snout - pelvic fin origin 262–293 | 275 | 7.5 | 247–285 | 271 | 11.1 | 254–304 | 278 | 15.7 |
| predorsal length 329–355 | 339 | 7.2 | 314–339 | 328 | 6.6 | 311–344 | 326 | 8.6 |
| pelvic fin origin - first dorsal fin 142–185 origin | 158 | 9.6 | 137–175 | 156 | 11.7 | 150–188 | 166 | 10.2 |
| body depth 124–169 | 141 | 12.8 | 112–167 | 141 | 15.4 | 134–181 | 155 | 12.1 |
| pectoral fin length 114–205 | 167 | 23.1 | 141–215 | 185 | 20.0 | 133–216 | 182 | 17.0 |
| first dorsal fin origin - anal fin 308–359 origin | 333 | 11.1 | 309–353 | 328 | 9.7 | 328–357 | 342 | 7.2 |
| pelvic fin origin - second dorsal 317–434 fin origin | 393 | 21.7 | 382–412 | 399 | 8.1 | 378–416 | 396 | 12.0 |
| first dorsal fin base length 219–317 | 260 | 22.8 | 219–331 | 266 | 25.5 | 210–321 | 272 | 27.7 |
| pelvic fin origin - anal fin origin 320–398 | 354 | 17.2 | 323–371 | 343 | 12.5 | 330–367 | 347 | 9.3 |
| second dorsal fin base length 115–153 | 134 | 9.2 | 93–159 | 134 | 15.4 | 128–160 | 143 | 7.9 |
| second dorsal fin origin - anal fin 117–147 origin | 134 | 7.5 | 121–149 | 134 | 8.2 | 135–159 | 145 | 5.9 |
| second dorsal fin origin - anal fin 139–177 insertion | 155 | 8.9 | 136–171 | 158 | 7.6 | 154–189 | 168 | 9.2 |
| anal fin origin - second dorsal fin 168–214 insertion | 183 | 10.2 | 155–220 | 189 | 15.7 | 180–213 | 195 | 9.8 |
| anal fin base length 101–139 | 119 | 9.4 | 103–149 | 125 | 12.0 | 117–154 | 133 | 11.1 |
| second dorsal fin insertion - anal 87–103 fin insertion | 97 | 4.6 | 94–119 | 101 | 6.3 | 91–108 | 98 | 3.6 |
| second dorsal fin insertion - cau- 197–251 dal fin | 231 | 13.6 | 203–270 | 236 | 16.2 | 205–239 | 228 | 8.2 |
| anal fin insertion - caudal fin 236–291 | 262 | 13.5 | 245–292 | 269 | 10.3 | 239–288 | 258 | 11.8 |
| caudal peduncle depth 65–79 | 72 | 3.4 | 72–84 | 76 | 3.5 | 70–79 | 74 | 2.6 |
TABLE 12. Sheared principal component loadings for morphometric variables for three new Percina species from the Mobile Basin. Measurements represent linear distances between landmarks as enumerated in Figure 1.
| Landmarks | Measurement | Size | Sheared PC II | Sheared PC III |
|---|---|---|---|---|
| 1 - 10 | standard length | 0.194 | 0.134 | -0.031 |
| 1 - 2 | snout length | 0.240 | 0.514 | 0.028 |
| 2 - 3 | eye diameter | 0.153 | 0.110 | 0.207 |
| 1 - 4 | head length | 0.198 | 0.254 | 0.087 |
| 5 - 16 | head depth | 0.217 | -0.071 | 0.061 |
| 5 - 15 | occiput - pelvic fin origin | 0.235 | -0.125 | 0.272 |
| 1 - 15 | snout - pelvic fin origin | 0.197 | 0.191 | 0.135 |
| 1 - 6 | predorsal length | 0.200 | 0.215 | 0.096 |
| 6 - 15 | pelvic fin origin - first dorsal fin origin | 0.214 | -0.147 | 0.333 |
| 6 - 14 | body depth | 0.224 | -0.280 | 0.459 |
| 17 - 18 | pectoral fin length | 0.141 | -0.138 | 0.324 |
| 6 - 13 | first dorsal fin origin - anal fin origin | 0.210 | 0.040 | 0.083 |
| 7 - 15 | pelvic fin origin - second dorsal fin origin | 0.185 | 0.061 | 0.053 |
| 6 - 7 | first dorsal fin base length | 0.219 | -0.141 | -0.192 |
| 13 - 15 | pelvic fin origin - anal fin origin | 0.195 | 0.130 | 0.105 |
| 7 - 8 | second dorsal fin base length | 0.233 | -0.169 | -0.276 |
| 7 - 13 | second dorsal fin origin - anal fin origin | 0.237 | -0.144 | -0.015 |
| 7 - 12 | second dorsal fin origin - anal fin insertion | 0.209 | -0.192 | -0.156 |
| 8 - 13 | anal fin origin - second dorsal fin insertion | 0.216 | -0.153 | -0.243 |
| 12 - 13 | anal fin base length | 0.200 | -0.355 | -0.287 |
| 8 - 12 | second dorsal fin insertion - anal fin insertion | 0.200 | 0.084 | -0.169 |
| 8 - 10 | second dorsal fin insertion - caudal fin | 0.181 | 0.252 | -0.173 |
| 10 - 12 | anal fin insertion - caudal fin | 0.190 | 0.261 | -0.180 |
| 9 - 11 | caudal peduncle depth | 0.178 | -0.007 | -0.155 |
TABLE 13. Analysis of covariance (ANCOVA) results for differences in log body depth between three new species of Percina from the Mobile Basin. Differences between species and the covariate (log SL) are significant; the interaction term tests for differences in slopes of the regression lines indicated in Figure 6 and is not significant. Sums of squares (SS) are Type III.
| Source df | SS | F | P |
|---|---|---|---|
| species 2 | 0.024 | 7.563 | 0.001 |
| log SL 1 | 0.120 | 74.427 | <0.001 |
| species*log SL 2 | 0.002 | 0.538 | 0.587 |
| Error 58 | 0.093 |
TABLE 14. Estimates of genetic diversity in the three new species described herein.
| N | Gene diversity | Nucleotide diversity Mean pairwise difference | Theta st |
|---|---|---|---|
| Percina sipsi 4 | 1.0 +/- 0.18 | 0.002 2.33 +/- 1.59% | 2.18 |
| Percina smithvanizi 6 | 1.0 +/- 0.09 | 0.003 3.53 +/- 2.09% | 4.38 |
| Percina kusha 5 | 0.7 +/- 0.22 | 0.003 4.00 +/- 2.40% | 3.36 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |