
Meru phyllisae, Spangler & Steiner, 2005
|
publication ID |
https://doi.org/10.1111/j.1365-3113.2005.00288.x |
|
DOI |
https://doi.org/10.5281/zenodo.4328176 |
|
persistent identifier |
https://treatment.plazi.org/id/03C5879C-FFC2-FF91-FC96-FDF1B8B4FC08 |
|
treatment provided by |
Plazi |
|
scientific name |
Meru phyllisae |
| status |
sp. nov. |
Meru phyllisae sp.n.
Description. Holotype, male, length 0.85 mm; width 0.39 mm. Wing condition undetermined. Other characters are as described for the genus.
Etymology. The specific name honours Phyllis M. Spangler who assisted in the discovery of this beetle and has been of great service to the study of aquatic Coleoptera for many years. without associated punctures (unlike others elsewhere on body); apical (sixth) sternite wider than long, apical margin rounded; surface bearing an irregular transverse band of flattened and grooved setae and fewer hairlike setae ( Fig. 8B View Fig ).
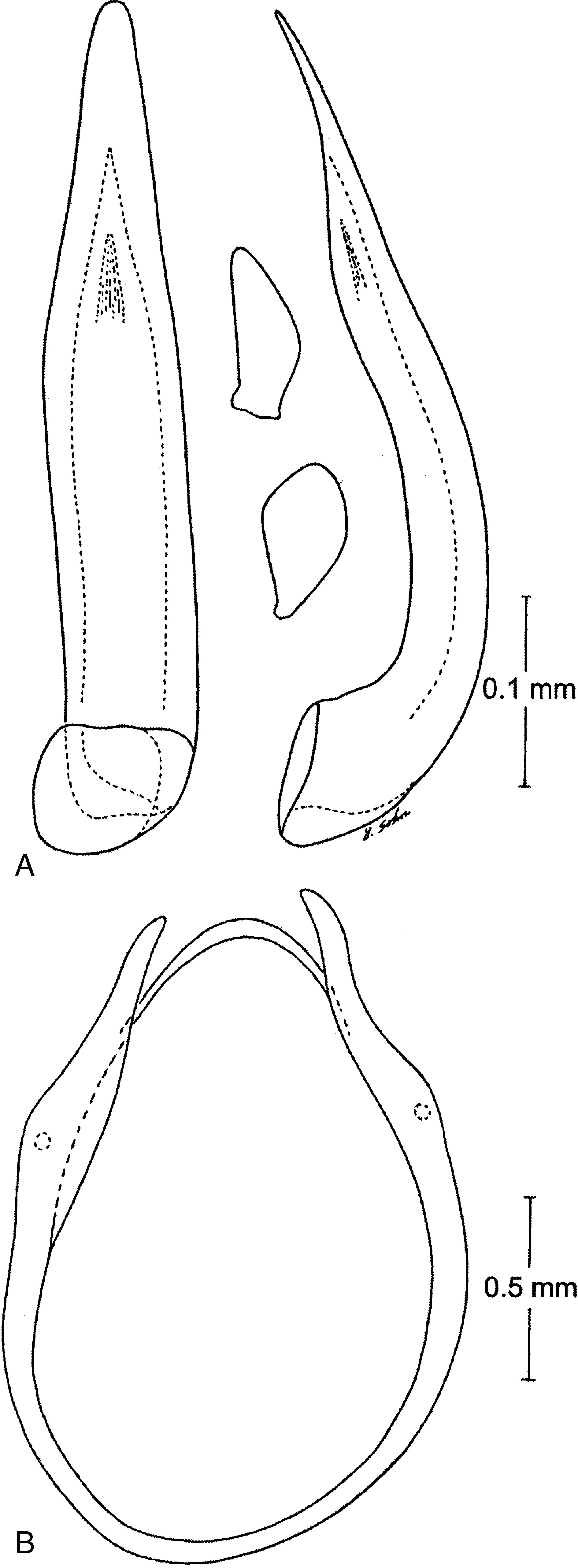
Male genitalia ( Fig. 14A View Fig ) asymmetric; basal piece absent (or fused with apical piece); median lobe with a curved, tubular base, becoming more straight, dorsoventrally flattened and tapered towards apex; parameres vestigial, scalelike, left paramere more rounded, laterally positioned at base of median lobe; right paramere smaller, more narrow, positioned on ventral side of base of median lobe. Aedeagus with only slight torsion in repose. Ring sclerite (ninth sternum) complete ( Fig. 14B View Fig ). Female genitalia lacking sclerotized structures except for reduced short gonocoxae, each bearing a few small apical setae ( Fig. 15 View Fig ).
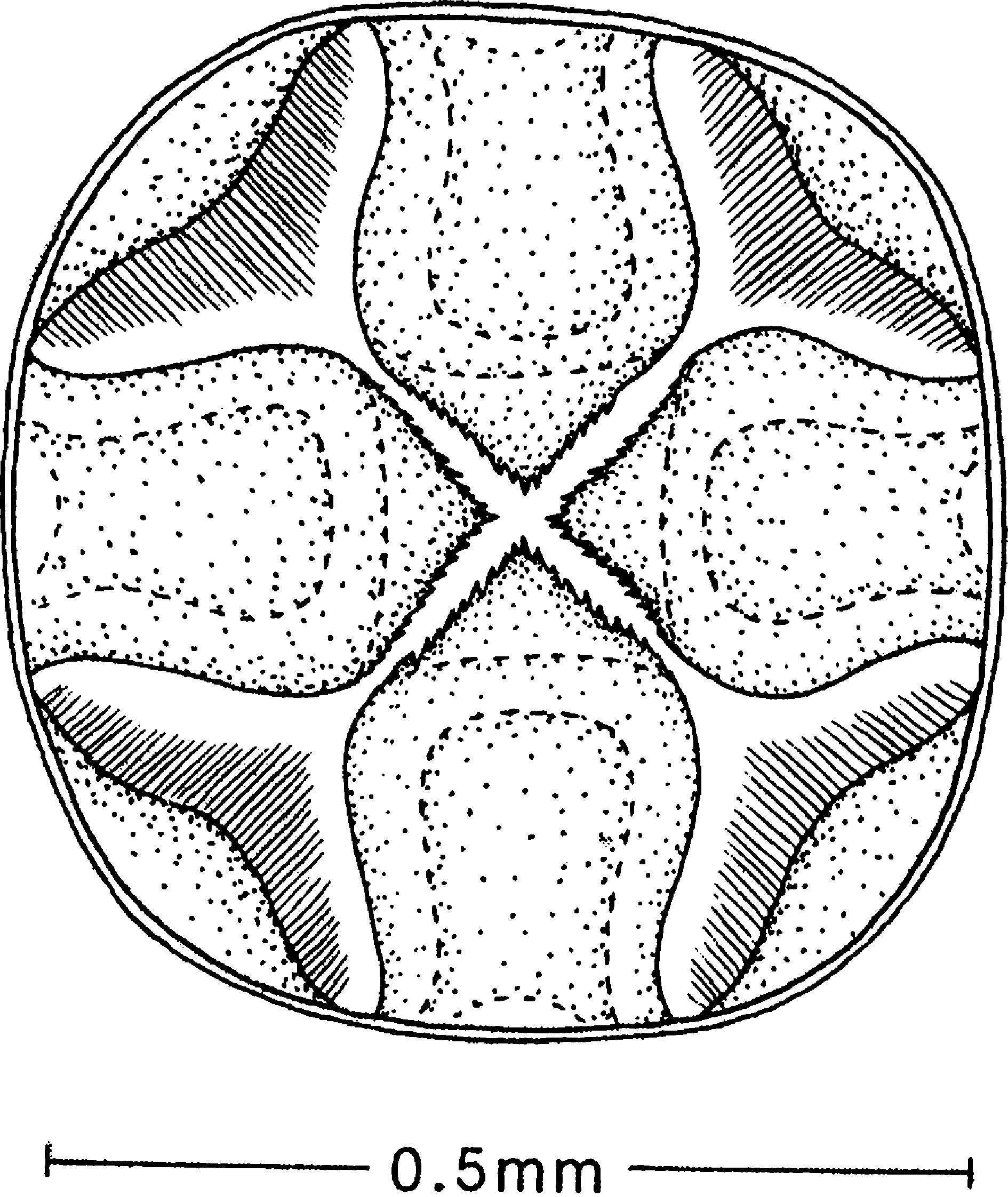
Proventriculus ( Fig. 16 View Fig ) with four larger plates smooth, each with single median acuminate and serrulate flap that appears to be capable of folding against the base; smaller four plates vase-shaped in side view, smooth, with a fringe of fine hairs.
Wing dimorphism. Of twelve specimens that were fully dissected, ten possessed atrophied wings and two had the fully developed condition.
Type data and material examined. Holotype?, labelled ‘ VENEZUELA: Amazonas, Puerto Ayacucho ( 40 km S) at Tobogan , 22 January 1985 /Seine of sunlit stream, slow water over sand & gravel, P. J. & P. M. Spangler, R. Faitoute, W. Steiner’ and 7 paratypes, same data. Other paratypes, 8, same data except ‘ 23 February 1986, sandy margins, P. J. Spangler, Colln. # 10’; 1, same data except ‘leaf packs, Colln. # 12, 24 February 1986, P. J. Spangler’; 1, same data except ‘ 26 February 1986, rootlets at shore edge, P. J. Spangler, Colln. # 19’; 7, same data except ‘ 27 February 1986, P. J. Spangler, Colln. # 20’; 19, same data except ‘ 28 February 1986, stream between cascades, P. J. Spangler, Colln. # 21’; 2, VENEZUELA, T. F. Amazonas, Puerto Ayacucho ( 40 km S) at Tobogań, 14 November 1987, Colln. # 1, P. J. Spangler & R. A. Faitoute’; 77, same data except ‘ 16 November 1987, Colln. # 7, P. J. Spangler & R. A. Faitoute’; 9, same data except ‘ 17 November 1987, Colln. # 11, P. J. Spangler & R. A. Faitoute’; 1, same data except ‘ 18 November 1987, Colln. # 13, P. J. Spangler & R. A. Faitoute //Collected by pouring water over stream bank & washing riparian insects into seine’; 3, ‘ VENEZUELA, T. F. Amazonas, Puerto Ayacucho ( 40 km S), El Tobogań, Caño Coromoto, 18 January 1989, P. J. Spangler, R. A. Faitoute & C. B. Barr //Collected at night with head lamp at upper shelter’; 2, same data except ‘ 20 January 1989, upper seep’; 1, same data except ‘ 20 January 1989, seined, at upper seep’; 1, same data except ‘ 23 January 1989, seined, stream between cascades’; 14, same data except ‘ January 1989 – reared’.
The holotype (and some paratypes from each of the labelled lots above) are deposited in the National Museum of Natural History , Smithsonian Institution, Washington, DC, U.S.A. ; other paratypes are deposited in the following institutions: The Natural History Museum , London ; Museum of Comparative Zoology , Harvard University , Cambridge ; California Academy of Sciences , San Francisco ; University of Arizona, Tucson ; Canadian National Collection , Ottawa ; Museo Argentino de Ciencias Naturales , Buenos Aires ; Instituto de Zoologia Agricola , Maracay ; Museum National d’Histoire Naturelle , Paris; Institut Royal des Sciences Naturelles de Belgique , Bruxelles; Naturhistorisches Museum, Wien; Zoologische Sammlung des Bayerischen Staates , Munich; Institut für Spezielle Zoologie und Evolutionsbiologie, Jena; Australian National Insect Collection, CSIRO, Canberra ; Entomological Laboratory, Kyushu University, Fukuoka, Japan .
Type locality. VENEZUELA: District of Amazonas, at El
Tobogań ; 40 km south of Puerto Ayacucho, 2 14 0 N 63 45 0 W GoogleMaps .
Habitats and collecting at the type locality
Various aquatic microhabitats were sampled at El Tobogań and yielded a rich assemblage of water beetle species. The cascade over the open bedrock ( Figs 17A, B, E, H View Fig ) was fed by an attractive white-water stream, the Rio (Caño) Coromoto, which originated in the mountains above. The stream emerged from the densely shaded jungle and flowed rapidly downhill through the forest for several hundred metres before the gradient flattened out as it reached an opening at a moderately deep, natural pool where the stream divided into two branches. Upon leaving the pool, the main water course flowed rapidly into a well-worn channel ( Figs 17A, E View Fig ) along the base of an adjacent hill. By contrast, the second branch fanned out and covered the broad expanse of granite bedrock with a shallow ( 3–5 mm) cover of slowly flowing water ( Fig. 17C View Fig ) with fallen leaves adhering to the wet rock. This broad area outlined the uppermost level of the entire cascade, which consisted of an upper and lower level; each about equal in length but the upper level was much broader and had a moderate gradient. The bedrock was obviously scoured from eons of periodic flooding and occasionally had an uncharacteristic deep pothole in its midst. The rock cascade emptied into a small, natural pool somewhat altered in past years by a man-made cement dam ( Fig. 17E View Fig ). Below the dam, the stream changed to a nearly level and narrower stretch of 30–35 m as it flowed through a tree-shaded area ( Figs 17F, G View Fig ) to the crest of a lower cascade. The stream was about 3 m wide, 0.3 m deep, and flowing about 1–1.5 m s 1 when the photographs ( Fig. 17G, H View Fig ) were taken. The substrate between the upper and lower cascades was rocky and sandy before the water tumbled over the narrower, steeper, and faster flowing lower cascade. The lower level was accessible on one side over an area of scoured granite bedrock. The opposite side of the stream was essentially inaccessible because of the dense jungle, deep channel, steeper gradient, and extremely fast current. Consequently, our collecting was conducted primarily on the broad, shallow surface and margins of the upper level, to and including the slower flowing area between the two cascade levels. Water analysis of the Caño Coromoto provided the following data: pH 5; oxygen 12 p. p.m.; hardness, 0. The water temperature was 28 C when most of the specimens were collected.
The following aquatic beetle taxa were also collected in and along the matted seepage areas that drained into the upper level of the cascade: Dytiscidae : Copelatus Erichson , Laccodytes Reǵimbart , Desmopachria Babington , various genera of Bidessini; Noteridae : Notomicrus Sharp , and a new genus; Elmidae : Cylloepus Erichson , Gyrelmis Hinton , Heterelmis Sharp , Austrelmis Brown , Microcylloepus Hinton , Neoelmis Musgrave , Pagelmis Spangler , Pilielmis Hinton , Phanocerus Sharp , Hintonelmis Spangler , Stegoelmis Hinton , Stenhelmoides Grouvelle , Tyletelmis Hinton , Xenelmis Hinton ; Hydraenidae : Adelphydraena orchymonti Perkins , Hydraena Kugelann ; Hydrophilidae : Anacaena Thomson , Berosus Leach , Chaetarthria Stephens , Notionotus Spangler , Oocyclus Sharp , Phaenonotum Sharp ; Hydroscaphidae : Scaphydra Reichardt ; Sphaeriusidae : Sphaerius Waltl ; Torridincolidae : Hintonia Reichardt.
The majority of the elmids were obtained from the rocky and shaded stream before it emerged from the jungle or the shaded part that connected the upper and lower halves of the cascade. Other specimens, including some M. phyllisae , were taken by collecting the leaves containing beetles from the seepage areas into a fine-mesh dip-net or seine. Partly submerged root mats that held decaying leaves, sticks and other debris at the edges of the open or semishaded stream margins ( Figs 17D, F, G View Fig ) were the main source of the specimens of Meru .
Observations on captive beetles
Live specimens of M. phyllisae collected 24 January 1989 were brought back to the laboratory and placed in a large glass finger bowl along with some decaying leaves. Daily observations were made and, from time to time, a search for eggs was conducted. Unfortunately, no eggs were found and no larvae appeared. The beetles spent most of their time sitting still or crawling about on the dead leaves. They were not observed feeding unless it was what appeared to be grazing over the leaf surfaces. Very rarely, a beetle would walk off a leaf into deeper water and simply float, posterior end uppermost, to the water surface. After floating against the surface film, it would turn over so that its venter was facing the surface and walk along under the surface film, as has been observed in Hydraenidae and some small Hydrophilidae . To submerge, the beetles would turn over with their head directed downwards and awkwardly ‘kick’ (with an alternating leg motion as in walking) their way downwards until finding a foothold on the substrate; if a beetle stopped ‘kicking’ on the way down, it would immediately float back to the surface, then start another descent. While the beetles were moving about, a silvery bubblelike area seemed to be present under each elytron; this was difficult to confirm because of the tiny size of the beetles.
No mortality was noted until almost 4 months after their capture, when two dead beetles were found on 24 May. After that, the following were found dead: one, 12 June; five, 11 July; two, 15 August. The last two specimens lived in the finger bowl for 196 days.



| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
| CSIRO |
Australian National Fish Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |