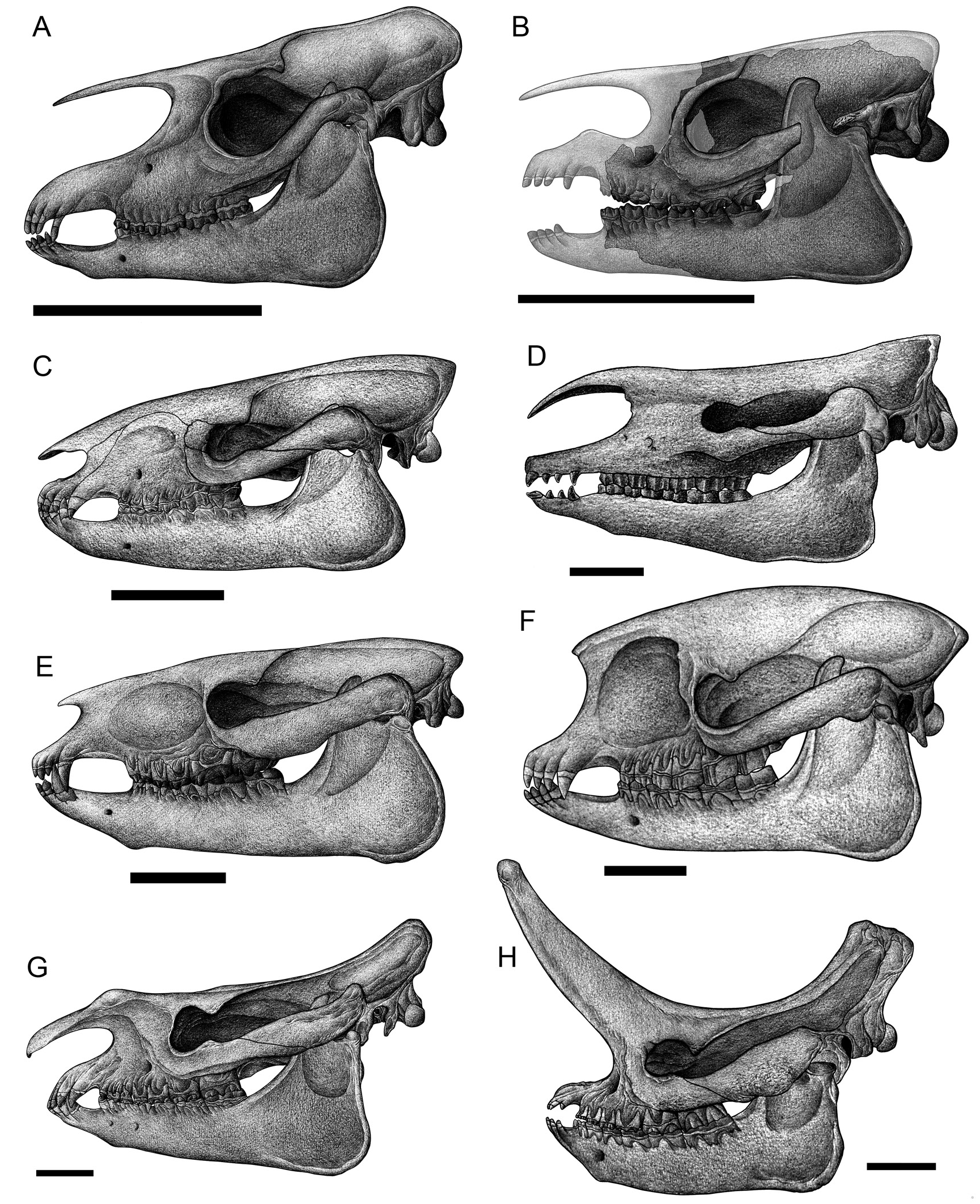
LOPHIALETIDAE MATTHEW AND GRANGER, 1925c
Minchenoletes Wang et al., 2011
Wang et al. (2011) erected a new genus and species,
Minchenoletes erlianensis
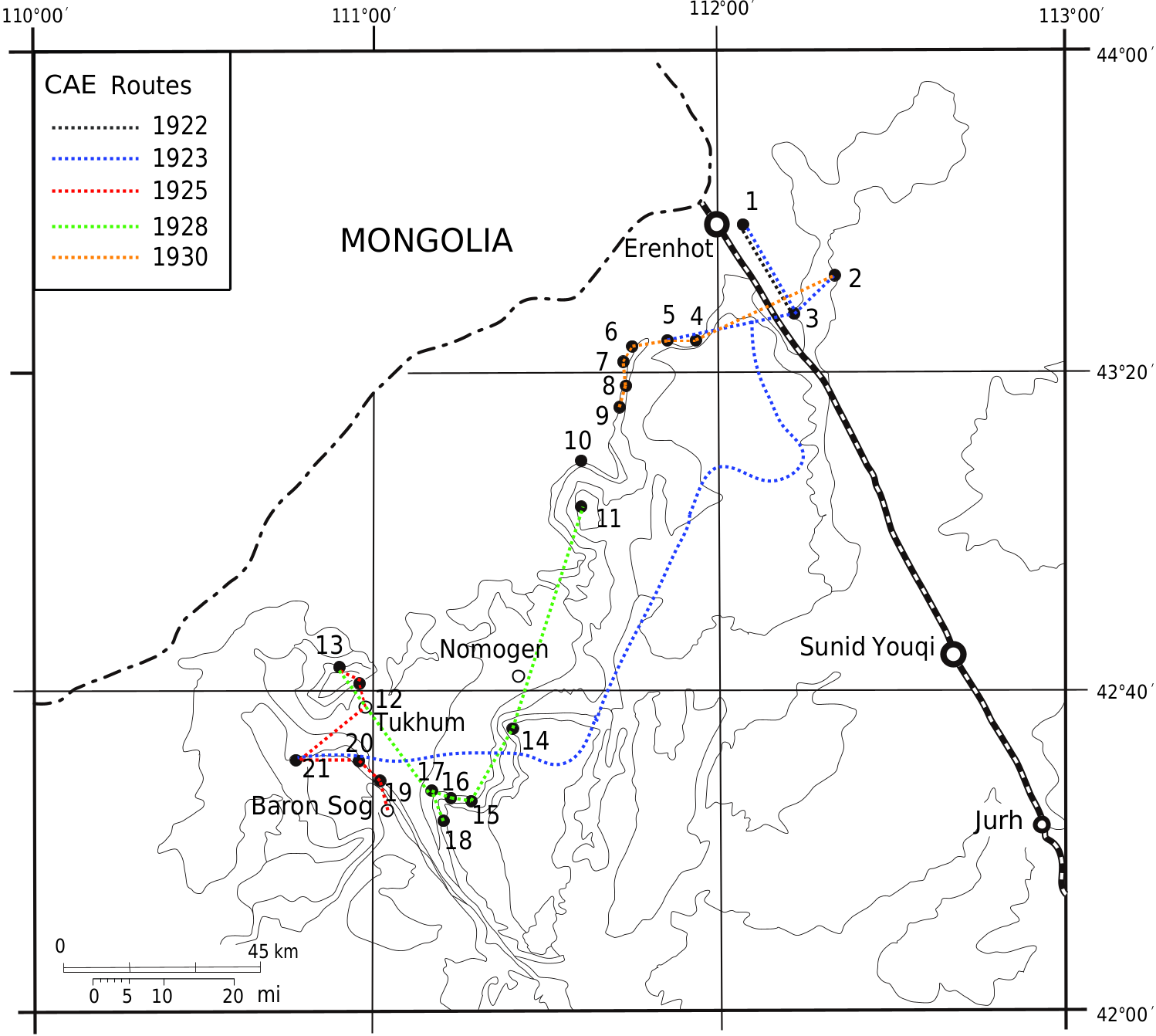
, from the upper part of the Nomogen Formation at Nuhetingboerhe and Wulanboerhe ( fig. 1
View FIG
). The specimens attributed to that taxon include maxillae, mandibles, and isolated teeth, and were unearthed from the NM-3 and 4 horizons of the Bumbanian ALMA.
Schlosseria Matthew and Granger, 1926
Only one species of
Schlosseria
,
S. magister
, has been named from the basin ( Matthew and Granger, 1926). The holotype (AMNH FM 20241) was unearthed from the Arshanto Formation, about 7 mi north of the Telegraph Line ( fig. 1
View FIG
). Radinsky (1965) assigned additional dental and postcranial material to
Schlosseria
cf.
S. magister
from Huheboerhe area. Those localities include Duheminboerhe, Daoteyin Obo, Huheboerhe, and Chaganboerhe. Our recent work confirms the presence of
Schlosseria magister
at Huheboerhe ( Li and Wang, 2010), which ranges from As-1 to As-6 fossil-bearing horizons and was considered restricted to the Arshanto Formation. However, Radinsky (1965) also considered AMNH FM 81787 and 81788 as
Schlosseria
cf.
S. magister
from “?Houldjin gravels” (i.e., Irdin Manha Formation) at Chaganboerhe.
Radinsky (1965) reported a lower jaw with p2–m2 of?
Schlosseria
(AMNH FM 26139) from “?Shara Murun beds” at Erden Obo (Urtyn Obo), and the fossil-bearing bed is probably equivalent to the “Basal White.” Li and Wang (2010) described a partial skull of
S. magister
(IVPP
V 16573
View Materials
) from the “Basal White” of Erden Obo. Our recent fieldwork also recovered some
Schlosseria
specimens from the basal layers at Wulanhuxiu, indicating the presence of Arshantan deposits in the section.
Lophialetes Matthew and Granger, 1925c
Only one species of
Lophialetes
,
L. expeditus
, is known from the Erlian Basin ( fig. 2A
View FIG
) ( Matthew and Granger, 1925c). The holotype (AMNH FM 19163) was found in the Irdin Manha Formation on the Irdin Manha escarpment. Radinsky (1965) also reported the species from both the “Irdin Manha” (i.e., Arshanto) and “Houldjin” (i.e., Irdin Manha) beds in the Huheboerhe area. However, he considered all these specimens as likely deriving from a single horizon. This proposal was supported by Li and Wang (2010) and Wang et al. (2010), who restricted
L. expeditus
to the IM-1 and IM-2 horizons of the Irdin Manha Formation.
Radinsky (1965) also reported
Lophialetes expeditus
? from the Ulan Shireh Formation at Wulantaolegai and Wulanhuxiu. These specimens are slightly smaller than Irdin Manha individuals of
L. expeditus
. He further assigned several larger specimens (AMNH FM 81687, 81690, and 81681) to
Lophialetes sp.
from the same bed at Wulanhuxiu ( Radinsky, 1965). Those specimens are approximately 20% to 25% larger than specimens of
L. expeditus
? from the same bed, and ~12% larger than Irdin Manha fossils of
L. expeditus
.
Radinsky (1965) assigned several crushed skulls, jaws, and foot bones (AMNH FM 22091– 22095) to
Lophialetes expeditus
? from the Tukhum Formation, which is overlain by the Shara Murun Formation at Ula Usu. He considered these specimens the same as Ulan Shireh
L. expeditus
? individuals. A lower jaw with p2–m2 (AMNH FM 26138) from “?Shara Murun beds” at Erden Obo was referred to
Lophialetes sp.
, which corresponds in size to the larger Ulan Shireh
Lophialetes
fossils ( Radinsky, 1965).
Zhongjianoletes Ye, 1983
Only one species of
Zhongjianoletes
,
Z. chowi
, known from a left mandible with a partial symphysis (IVPP V 6671), is known from the Ulan Shireh Formation at North Mesa ( Ye, 1983). The species is very similar to
Lophialetes expeditus
, but is larger and lacks the p1 and probably incisors. Ye (1983) further considered that specimens referred to
Lophialetes sp.
by Radinsky (1965) from the same locality should belong to
Z. chowi
. Ye (1983) assigned some isolated M3s to
Zhongjianoletes sp.
, which corresponds to the size of
Z. chowi
from the same horizon.
Simplaletes Qi, 1980
Two species of
Simplaletes
,
S. sujiensis
and
S. ulanshirehensis
, are known from the Irdin Manha and Ulan Shireh, respectively ( Qi, 1980).
Simplaletes sujiensis
was found in the Irdin Manha Formation, and
S. ulanshirehensis
was discovered in the Ulan Shireh Formation in 1959 by SSPE. Both of those species preserve only lower jaws with relatively heavily worn teeth, and lack the p1. Lucas et al. (1997) considered
Simplaletes
to be a junior synonym of
Schlosseria
.
Breviodon Radinsky, 1965
Two species of
Breviodon
,
B. acares
and B.?
minutus
, are known from the Wulanhuxiu and Irdin Manha, respectively ( Radinsky, 1965). The holotype of
B. acares
is a mandible (AMNH FM 26113) characterized by the absence of p1–2 and it is about 30% smaller than Ulan Shireh material of
Lophialetes expeditus
?. Radinsky (1965) assigned a fragmentary skull (AMNH FM 81751) to
Breviodon
cf.
B. acares
from the “Irdin Manhan beds” at Huheboerhe. However, this specimen more likely was recovered from the Arshanto Formation instead of the “Irdin Manha Formation.” A lower jaw with dp4–m2 (AMNH FM 81836) from the same locality may be conspecific with the fragmentary skull ( Radinsky, 1965). Reshetov (1975) erected a new genus
Parabreviodon
based on AMNH FM 81751 as the generic type. A lower jaw (AMNH FM 26115, field no. 645) from Wulanhuxiu and a few isolated teeth (AMNH FM 81839) from the “Irdin Manha beds” (i.e., the Arshanto Formation) at Daoteyin Obo, respectively, were assigned to?
Breviodon
based on inadequate material ( Radinsky, 1965).
Radinsky (1965) reassigned the holotype of “
Lophialetes
”
minutus
(an isolated left upper molar, AMNH FM 20139) and some miscellaneous teeth (AMNH FM 81721A, 81721B, 81722) from the Irdin Manha Formation of the Irdin Manha escarpment to
Breviodon
?
minutus
. However, Reshetov (1979) considered
Breviodon acares
as a junior synonym of
B. minutus
based on the associated upper and lower cheek teeth. This notion was followed by Qi (1987), who also reported the species from the Arshanto Formation at Huheboerhe and Wulanboerhe.
Rhodopagus Radinsky, 1965
Two species of
Rhodopagus
,
R. pygmaeus
and R.?
minimus
, are known from North Mesa and Ula Usu, respectively ( Radinsky, 1965).
Rhodopagus pygmaeus
is known mainly from the Ulan Shireh Formation, and Radinsky (1965) also assigned a couple of specimens (AMNH FM 81842, 81843) to?
R. pygmaeus
from the type Irdin Manha bed. Radinsky (1965) transferred
Caenolophus
?
minimus
(AMNH FM 20310) from the Shara Murun Formation at Ula Usu to
Rhodopagus
?
minimus
based on its short trigonid and reduced cristid obliqua. Lucas and Schoch (1981) further considered that
Rhodopagus pygmaeus
is a junior synonym of
R. minimus
.
The phylogenetic position of
Rhodopagus
is controversial with its affinities to
Lophialetidae
,
Helaletidae
, or even
Hyracodontidae (Lucas and Schoch, 1981)
. Our recent discovery of
Rhodopagus
from the “Basal White” of Erden Obo will probably clarify this phylogenetic mystery in the future.
Pataecops Radinsky, 1966
Only one species of
Pataecops
,
P. parvus
, is known from the upper part of the Nomogen Formation at Nuhetingboerhe and Wulanboerhe ( Wang et al., 2011). Wang et al. (2011) pointed out that the holotype of the species from the Kholobolchi Formation of Mongolia is Bumbanian in age, as are specimens from the Erlian Basin. Although Radinsky (1965) assigned
Pataecops
to
Lophialetidae
, the phylogenetic position of
Pataecops parvus
is uncertain. If
P.parvus
belongs to rhinocerotoids rather than tapiroids (Lucas and Schoch, 1981; Dashzeveg and Hooker, 1997), it would extend the earliest record of rhinocerotoids into the earliest Eocene ( Wang et al., 2011).