
Triacanthoneus akumalensis, Alvarez, Fernando, Iliffe, Thomas M., Gonzalez, Brett & Villalobos, José Luis, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.279609 |
|
DOI |
https://doi.org/10.5281/zenodo.6178317 |
|
persistent identifier |
https://treatment.plazi.org/id/03C33854-FFDA-710A-FF18-FF2B324BFDDC |
|
treatment provided by |
Plazi |
|
scientific name |
Triacanthoneus akumalensis |
| status |
sp. nov. |
Triacanthoneus akumalensis View in CoL sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Type material. Holotype: ovigerous female, cl 8.9 mm, tl 26 mm, Mexico, Quintana Roo, Akumal, Cenote Aak Kimin, underwater cave (20o 24’ N, 87o 18’ W), depth 25–40 m, 15 April 2008, coll. T.M. Iliffe, CNCR 26315. Paratypes: 1 non-ovigerous specimen (male?), cl 9.1 mm, tl 25.9 mm, same data as for the holotype, CNCR 26316; 1 ovigerous specimen with appendix masculina, cl 8 mm, tl 23.7 mm, same data as for the holotype, CNCR 26486.
Description. Carapace broadly rounded laterally, with faint anterolateral suture and two strong, acute postorbital teeth; dorsal midline with prominent sharp tooth in epigastric position, low carina present between epigastric tooth and orbit; posterior margin with deep cardiac notch ( Figs. 1 View FIGURE 1 a, b). Rostrum reaching mid-length of third antennular article, tip slightly pointing upwards, ventral margin with small tooth ( Figs. 1 View FIGURE 1 b, 3a).
Eyes completely covered by carapace, not visible in dorsal view; lateral region pigmented, cornea not faceted; dorsal margin with small acute projection present ( Figs. 3 View FIGURE 3 a, f).
First three abdominal somites with lateroventral margins of pleura rounded; fourth somite with lateroventral margin subtriangular; fifth somite with lateroventral margin produced in a small acute tooth; sixth somite longer than other somites, without articulated plate.
Telson approximately 2.6 times as long as its anterior width; posterior margin with deep “V” shaped notch, each posterolateral angle with a pair of long spines, lateral longest; posterior half of dorsal surface with shallow median longitudinal groove and two pairs of short spines ( Fig. 3 View FIGURE 3 b). Uropods with both rami longer than telson, rounded distally; exopod longer than endopod, with complete diaeresis and strong distolateral spine.
Antennule with prominent stylocerite, its acute tip reaching beyond mid-length of second article ( Fig. 3 View FIGURE 3 a); second article longer than wide, about as long as third article; external flagellum with four basal articles, accessory branch with about eight articles bearing long aesthetascs ( Fig. 3 View FIGURE 3 c). Antenna with basicerite armed with stout, sharp distal tooth; carpocerite short, not reaching beyond half-length of scaphocerite; scaphocerite ovate, with rounded anterior margin reaching beyond distolateral tooth ( Fig. 3 View FIGURE 3 a).
Mouthparts typical for genus. Mandible with incisor process bearing nine teeth distally; molar process cylindrical, stout, with blunt distal surface; palp two-articulated, distal article oval-shaped ( Fig. 2 View FIGURE 2 a). Maxillule with bilobed palp, each bearing one long seta ( Fig. 2 View FIGURE 2 b). Maxilla with simple elongate palp, devoid of setae; distal endite bilobed, lobes divided by a deep cleft; scaphognathite about 3.2 times as long as wide ( Fig. 2 View FIGURE 2 c). First maxilliped with stout palp bearing distally two long setae covered with short setules; distal endite broad, subtrapezoidal; exopod elongate, caridean lobe evident; epipod approximately semicircular ( Fig. 2 View FIGURE 2 d). Second maxilliped with elongate exopod; endopod with ischium and dactylus of about the same length; epipod relatively broad, somewhat elongate, as long as ischium of endopod; podobranch-like gill well-developed ( Fig. 2 View FIGURE 2 e).
Third maxilliped pediform, slender; endopod three-articulated, antepenultimate article subequal in length to penultimate and ultimate articles combined; ultimate article longer than penultimate, mesial surface covered with rows of short setae, tip with tuft of long setae; exopod shorter than antepenultimate article of endopod; epipod present as mastigobranch emerging from base of rounded lateral plate; arthrobranch well-developed ( Fig. 2 View FIGURE 2 f).
First pair of pereopods asymmetrical, major cheliped always on the right side ( Fig. 1 View FIGURE 1 a). Major cheliped 0.75 times of total body length; ischium about one third of merus length, with three ventral spines; merus with ventral margin depressed, dorsal margin straight, distally somewhat thickened; carpus shorter than merus, wider distally; propodus strongly arched; palm about 2.4 times as long as high, inflated, with low hump on dorsal surface, adjacent to articulation with dactylus; fingers about 1.3 times length of palm, strongly crossing distally; cutting edge of fingers serrated with minute teeth, and with four-five conspicuously larger teeth intercalated among smaller teeth ( Fig. 1 View FIGURE 1 h). Minor cheliped approximately half as long as major cheliped, much less stout (fig. 1a); ischium with three ventral spines; ischium, merus, carpus and propodus subequal in length; chela simple, dactylus slightly shorter than palm ( Fig. 3 View FIGURE 3 g).
Second pair of pereopods slender ( Fig. 1 View FIGURE 1 b, 3i); ischium and merus subequal in length; ischium with row of three spines on ventrolateral surface; carpus five-articulated, first carpal article longest, more than half-length of the entire carpus, remaining carpal articles of about the same length; chela simple, with fingers longer than palm. Third to fifth pairs of pereopods slender with simple dactyli, increasing in length posteriorly ( Fig. 1 View FIGURE 1 b); third and fourth pereopods each with three spines on ventrolateral surface of ischium; carpus longest article in third and fourth pereopods; propodus longest article in fifth pereopod.
Pleural process, strong and anteriorly arched, arising from lateral body wall, above coxa of fifth pereopod ( Fig. 1 View FIGURE 1 c).
Second pair of pleopods of male paratype with protopod bearing long thick setae curling distally ( Fig. 3 View FIGURE 3 d); exopod longer than endopod; endopod with appendix interna and appendix masculine, former elongate, slightly longer than appendix masculina; appendix masculina cylindrical, with nine strong spines along lateral and distal margins ( Fig. 3 View FIGURE 3 e).
Maxillipeds Pereopods
1rst 2nd 3rd 1rst 2nd 3rd 4rth 5th Pleurobranch - - - P P P P - Arthrobranch - - P - - - - - Podobranch - P - - - - - - Mastigobranch - P P P P P P - Exopod P P P - - - - - Etymology. The specific epithet refers to the type locality, Akumal, Quintana Roo, Mexico.
Distribution. Presently known only from the type locality, Cenote Aak Kimin, Akumal, Quintana Roo, Mexico’s Yucatan Peninsula (see below).
Habitat. Cenote Aak Kimin is located 100 m inland from the Caribbean coast at Half Moon Bay, about 1.5 km northeast of Akumal and 0.5 km southwest of Yal-ku Lagoon. While most cave passages in the Yucatan Peninsula are primarily oriented perpendicular to the coast and act as subterranean rivers, this 2 km long cave runs parallel to and just inland from the shoreline under a mangrove swamp. The cave entrance is at a depth of 3 m in a small (7–8 m diameter) brackish pool surrounded by red mangrove forest. The cave has two distinctly different sections with discrete modes of formation. The upper, shallow water (6 m) passages were created by solutional processes beneath the mangrove swamp and form a network of low but wide tunnels extending parallel to the coast and connecting through to the Yal-ku Lagoon. The lower deep section of the cave runs beneath the shallow passages and consists of a narrow fissure with vertical walls. It was formed from slump faulting on the platform margin and enlarged by solutional processes. Small stalactites and stalagmites are present throughout the shallow section but are brittle to the touch due to mixing corrosion between fresh and salt water. Water in the shallow water section of the cave is regulated by tidal pumping and contains a mix of both stygobitic and ocean species. Tidal pumping does not appear to be a factor within the deeper section of the cave. This section is more biologically depauperate, with only stygobites observed, including the shrimps, Typhlatya sp. and Yagerocaris cozumel Kensley, 1988 , and the isopod, Metacirolana sp. The specimens of Triacanthoneus akumalensis sp. nov. were collected at depths between 25 m and 40 m.
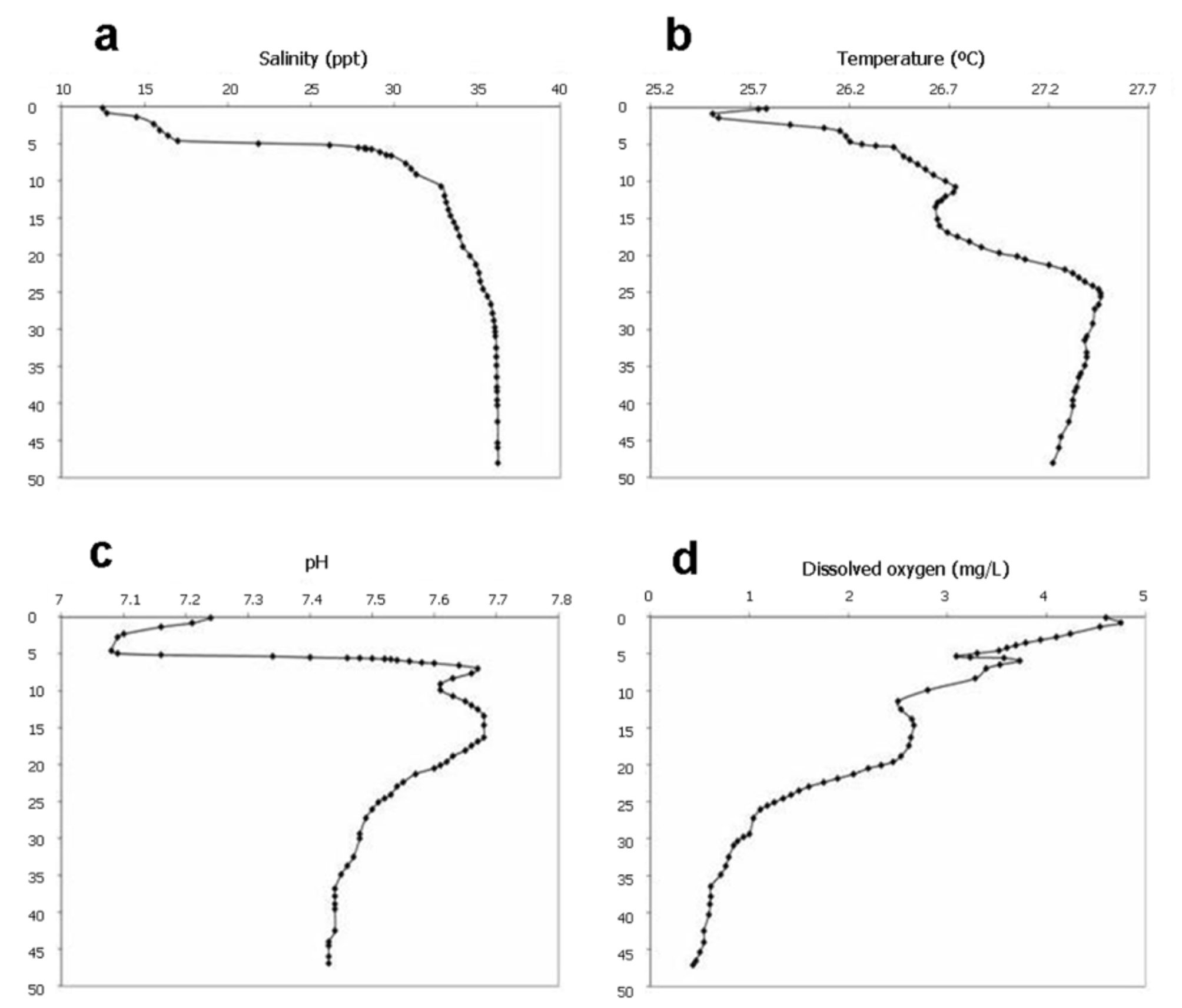
A water column profile in Cenote Aak Kimin was obtained in March 2011 by cave divers using a YSI 600 XLM Multiparameter Water Quality Analyzer held forwards during the descent. Surface waters of the entrance pool were brackish ( Fig. 4 View FIGURE 4 a). A halocline was present between 4.9 and 9.3 m marking the transition to marine waters (32.8 ppt) with salinity increasing to 36.26 ppt at depths below 30 m. While insolation warms the surface pool (25.83 °C), a small drop in temperature occurred at slightly deeper depths, followed by a gradual increase across the halocline ( Fig. 4 View FIGURE 4 b). A second noticeable temperature increase at 10.7 m is likely associated with tidal pumping from the nearby Yal-ku Lagoon. A temperature maximum (27.51 °C) at 25 m with was followed by a gradual decrease in temperature with depth, possibly associated with deep water circulation. A pH minimum (7.08) occurred at the top of the halocline, followed by two maxima at 7 and 15 m, with a gradual decrease then to 7.43 at maximum depth ( Fig. 4 View FIGURE 4 c). Dissolved oxygen at the surface of the entrance pool was 4.75 mg /L with a slight decrease at the halocline ( Fig. 4 View FIGURE 4 d). A small rebound in dissolved oxygen was observed at 15 m, probably due to horizontal water flow of tidally pumped water from nearby Yal-ku Lagoon. The dissolved oxygen concentration decreased to a low of 0.43 mg /L at 47 m. Redox levels were distinctly bimodal (data not shown), suggesting significant reducing conditions below the halocline and throughout the deep sections of the cave.
Remarks. The new species differs from the other three species of Triacanthoneus in several respects. The eyes of T. akumalensis sp. nov. are completely covered by the carapace and are not visible in dorsal view as they are in the other three species. The rostrum, as seen in the dorsal view, is elongate-triangular in the three previously known species, while it is more acuminate in the new species. In the three previously described species, the dentition of the major chela is formed by similar-sized teeth, alternating on both fingers, whilst in the new species, four or five large teeth on each finger are separated by regular spaces themselves bearing smaller teeth. The posterior margin of the telson has a deep notch in the new species; it is straight in the other three species. In addition, the rostra of T. toro and T. pacificus are shorter than in T. akumalensis , not reaching beyond the second article of the antennular peduncle, whilst T. alacranes has orbital teeth, which are absent from the other three species, including T. akumalensis sp. nov.
The branchial formula reported for the previously described species of Triacanthoneus ( Anker 2010) differs from that of T. akumalensis sp. nov. in lacking a pleurobranch on the fifth pereopod and having an arthrobranch on the second maxilliped. Furthermore, T. akumalensis sp. nov. has a fairly strong process above the coxa of the fifth pereopod, in the area of insertion of a pleurobranch, perhaps a modified pleurobranch; this character is presumably absent from the other three species.
Anker (2010) reported the presence of an appendix masculina in all individuals of Triacanthoneus , including ovigerous females, suggesting some sort of hermaphroditism. In T. akumalensis sp. nov., the ovigerous female holotype does not have an appendix masculina; the paratype specimen sexed as male does not show any evidence of egg incubation or maturation; however, the second paratype has both eggs and a well-developed appendix masculina. At this stage, with the evidence coming from only three individuals, we could suggest the presence of another variation of hermaphroditism in T. akumalensis . Another difference between the previously described marine species of Triacanthoneus and the cave-dwelling T. akumalensis sp. nov. is the size: the carapace length of T. akumalensis sp. nov. (8.0, 8.9, 9.1 mm) is more than twice to three times that of the three previously described species (maximum cl 2.8, 3.7, 4.1 mm).
In respect to the anchialine caridean fauna of the Yucatan Peninsula, Triacanthoneus represents the eleventh caridean genus and the second alpheid genus after Yagerocaris Kensley, 1988 to be recorded in this area ( Kensley 1988; Anker 2008). In addition, T akumalensis becomes the 14th species of anchialine shrimp in Yucatan ( Alvarez & Iliffe 2008). Within the family Alpheidae , T. akumalensis sp. nov. is the 10th described species reported from cave and anchialine habitats, with several more awaiting description (see Anker 2008).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |