
Costasiella coronata, Swennen, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.4509064 |
|
persistent identifier |
https://treatment.plazi.org/id/03C2AB0B-9639-F27C-DAE7-33224A1B35F1 |
|
treatment provided by |
Carolina |
|
scientific name |
Costasiella coronata |
| status |
sp. nov. |
Costasiella coronata , new species
( Figs. 1 View Fig , 2 View Fig )
Material examined. – Bang Tawa, in the mangrove forest (6°51'N 101°09'E), some specimens and egg-strings, 10 Jul.2000. Same site, some tens of individuals with a length up to 7 mm, also egg-strings found, Oct.2000. Same site, some tens of individuals with a length of 4-10.5 mm, also egg-strings found, 13 Oct.2003. Tak Bay, on intertidal mud bordering a small fringe of mangroves (6°16'N 102°03'E), 2 specimens with a length of 4-5 mm, 23 Apr.2005. Both locations are in southern Thailand along the Gulf of Thailand, and all specimens were on, or in, patches of the alga Vaucheria sp GoogleMaps .
The description is based on 11 specimens collected in Oct.2003, of which five were dissected and six selected as types. The holotype and three paratypes are deposited in the Zoological Reference Collection ( ZRC) of the Raffles Museum of Biodiversity Research, National University of Singapore, and three paratypes in the Zoological Museum, University of Amsterdam ( ZMA).
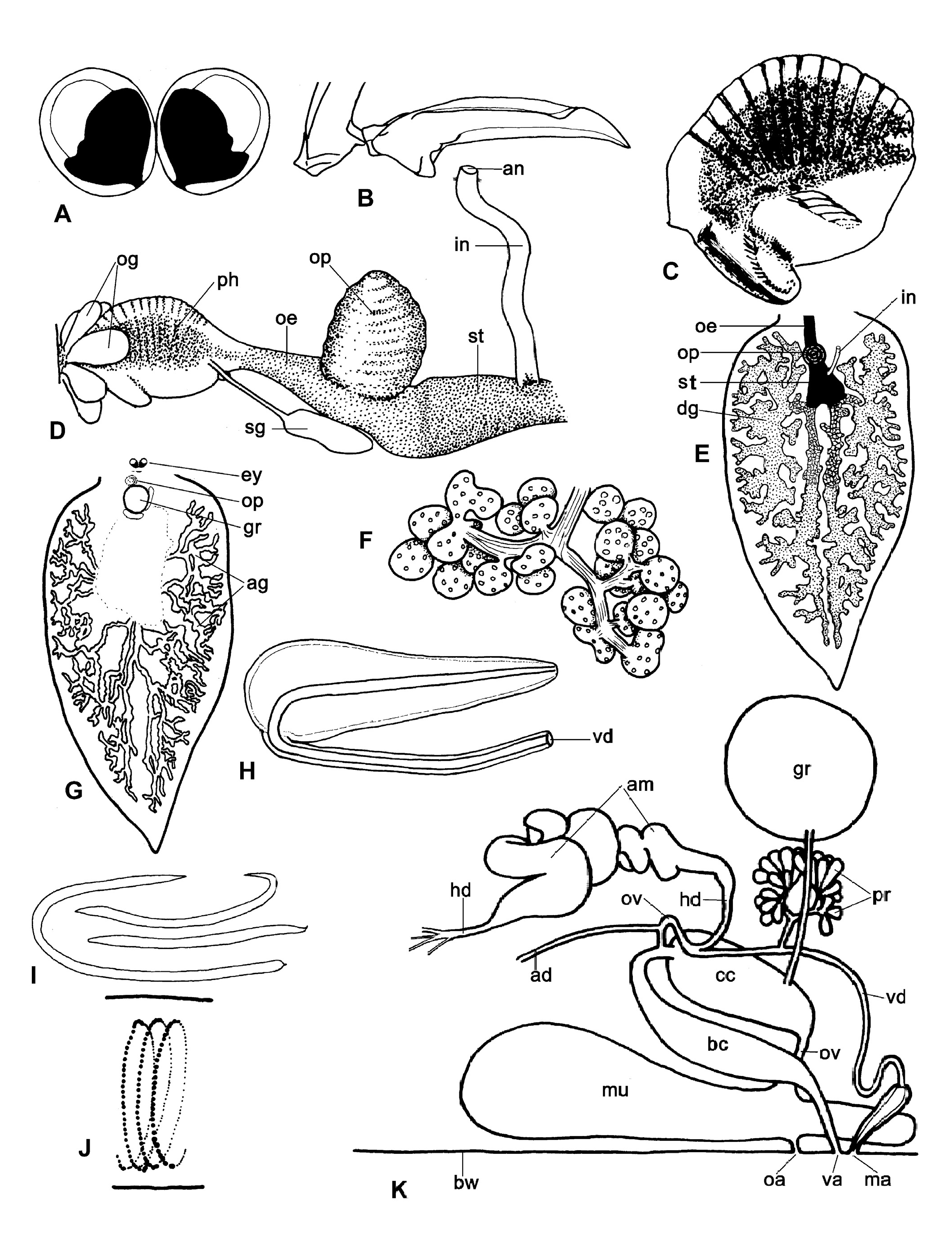
External morphology. – Black eyes close together between rhinophores ( Fig. 1 A, C View Fig ). Renal shield large, fixed on dorsal side of small pericardium, covering dorsum from just posterior of eyes to two-third of body length; wide, free rim can be held upwards and pleated ( Fig. 1 A View Fig ). Rim usually covers basal parts of bordering cerata and often nearly their whole length ( Fig. 1 B View Fig ). Transparent renal shield contains irregular central channel with side-branches bordered by dispersed dark dots. Rhinophores smooth, wide, somewhat flattened at base, but rounded distally and tapering into blunt tips. Shape of head varies according to degree of straightening ( Fig. 1 C View Fig ); frontal part usually kept bilobed. Lateral areas of back wider than body by thick rims ( Fig. 1 D, E View Fig ). Each rim filled with maze of branches of albumen and digestive glands. Cerata in a row of up to 22 on dorsal side of each rim, starting shortly behind rhinophores; not covering central part of dorsum and posterior end of foot; strongly flattened, wide, and smooth ( Fig. 1 F, G View Fig ); flat sides concave-convex in preserved specimens ( Fig. 1 G View Fig ). Each ceras containing wide branch with side branches of digestive gland ( Fig. 1 F View Fig ) and in reproductive slugs also branches of albumen gland being most dense on dorsal side of ceras ( Fig. 1 H View Fig ).
Foot somewhat wider than dorsum in live specimens, but the opposite is true after preservation. Anterior border of foot with blunt extensions; posterior tip pointed ( Fig. 1 A, E View Fig ). Foot length 4-6 times width in actively crawling specimens (N = 11). Sole anteriorly with shallow central longitudinal groove continuing posteriorly as unpigmented line. Anus on small papilla anterior right of pericardium covered by renal shield ( Fig. 1 I View Fig ). Renal pore just left of anus papilla. Male aperture below right rhinophore, vaginal aperture just posterior of male aperture and oviductal aperture slightly posterior to and below vaginal aperture ( Fig. 1 J View Fig ).
Live colouration. – Colour of body and cerata is mainly determined by colour of digestive gland: bright green in well-fed individuals, and pale yellowish brown when not fed recently. Skin largely transparent with scattered iridescent blue-green dots, white or yellowish glandular dots, and angular reddish-brown dots, latter especially on renal shield. These colours only visible under specific light conditions. Moreover, fine grey-purple pigment on lateral and dorsal sides, anterior foot border, and in two longitudinal bands on sole of foot. White lines on posterior sides of rhinophores and thin dark pigment on lateral sides. Appearance of lines and areas with grey-purple pigment on head largely depends on degree of expansion of head ( Fig. 1 C, I View Fig ). A small black line present on both sides of mouth ( Fig. 1 K View Fig ). No pigment around genital orifices and eyes. In preserved specimens content of digestive gland brownish and purple areas in epidermis grey.
Internal characteristics. – Diameter of eyeball 125–145 µm (N = 4, length of live specimen 6–8 mm). Eyeballs touching ( Fig. 2 A View Fig ). Clusters of oral glands besides anterior part of pharynx ( Fig. 2 D View Fig ). Pharynx nearly spherical, diameter 400– 520 µm (N = 5) with prominent ascus, strong ascus muscle, and well developed dorsal septate muscle. Wide, internal, longitudinal band of dark pigment borders pharyngeal space ( Fig. 2 C, D View Fig ). Radular teeth blade-shaped, smooth and narrow, with slightly curved sharp tips ( Fig. 2 B View Fig ). Length of leading tooth in large specimens 85–100 µm (N = 5). In ascending row 5-8 teeth, in descending row and ascus 18–27 progressively smaller teeth (N = 5). Teeth, including 3 preradular teeth, remain fixed on radular ribbon that curls in a spiral in ascus. Length of base of first preradular tooth about 4 µm, second about 8 µm, third with minuscule blade 15 µm in total. On both sides of short oesophagus, salivary duct between pharynx and salivary gland below prominent oesophageal pouch on dorsal side of oesophagus just anterior of stomach ( Fig. 2 D View Fig ). Oesophagus and pouch dark pigmented; salivary glands and ducts pale. Stomach dark, ventral in body. From right dorsal side of stomach, pale coloured intestine ascends to anal papilla ( Fig. 2 D View Fig ). A branch of the digestive gland starts in both posterior corners of stomach. Both immediately split into a branch that goes slanting upwards and a branch that directly goes in posterior direction and then upwards (N = 5). Both branches knotty, repeatedly branching in respectively anterior and posterior dorsal parts of body. First branch reaches to just posterior of eyes, second up into tail far posterior of origins of posterior cerata ( Fig. 2 E View Fig ). Only some branches of digestive gland in dorsum connected with a ceras.
Extensive network of albumen gland tubules ramifying in dorsal side of body close to branches of digestive gland including in cerata ( Figs. 1 H View Fig , 2 G View Fig ). Tubules white and relatively thick in egg laying specimens, thin and pale in non-reproductive individuals, uniting into single albumen duct. Hermaphroditic follicles dense ( Fig. 2 F View Fig ), small (diameter single follicles 50-150 µm, length of compound follicles 200-300 µm), and numerous (>500), on ventral side of body cavity from near frontal border of foot to below division between anterior and posterior networks of digestive gland. Follicles yellow, connected by ductules that unite into short hermaphrodite duct. Organs of genital complex ( Fig. 2 K View Fig ) packed together in anterior right half of body cavity. Hermaphrodite duct widens into strongly coiled ampulla leaving it at anterior end and splitting into vas deferens and small oviduct. Vas deferens receives prostate duct coming from about 30 fusiform prostate glands in anterior part of body below spherical genital receptacle, then running anteriorly over female system to long, narrow, unarmed penis ( Fig. 2 H View Fig ). Male system uncoloured, but prostate glands white. Vaginal duct widens into bursa copulatrix going to posterior end of solid, ovoid central canal. Small oviduct receives albumen duct and enters central canal together with vaginal duct. Oviduct leaves central canal in anterior end and continues as large oviduct via wide and voluminous mucous gland occupying most of ventral right of anterior body cavity. Genital receptacle spherical, pale, situated dorsally slightly right of oesophageal pouch and left of loop of bursa copulatrix (vaginal duct). Genital receptacle connected to left side of central canal by narrow tube.
Habitat and behaviour. – Specimens of Costasiella coronata were found on, and shallowly buried into, soft muddy sediment in and around tufts of Vaucheria sp. in open spaces within or bordering mangrove forests high in the intertidal zone. The slugs lived amphibiously. When the Vaucheria patches had emerged during low tide, more and more slugs crawled over the algae for feeding. The contracted rhinophores were firmly kept against the back and the cerata were held largely under the renal shield that seemed to increase in thickness and relative size ( Fig. 1 B View Fig ). The epidermis looked dirty by mud suggesting that the ciliary actions were reduced or had ceased completely. The number of visible slugs declined again by hiding in mud when the tide was out for some time. In captivity it could be seen that they fed by sucking the siphonal cells of Vaucheria sp. both when submerged and when emerged. In October 2000 the temperature of the water was 35°C and the salinity 29‰, but salinity fluctuates strongly over the season in the coastal zone ( Swennen et al., 2001).
The slugs were difficult to keep alive in captivity for more than a few weeks. The main problem was maintaining the algae in a condition such that they could be consumed by the slugs; replacing the algae every few days was necessary. A good growth of the Vaucheria sp. did occur when keeping them with substrate either permanently emerged in a damp atmosphere, or submerged with some aeration, but in both cases the slugs either hardly fed or did not feed at all.
Copulation was not observed in individuals out of the water. When two submerged slugs met for mating, they erected their rhinophores and repeatedly touched each other with the swollen anterior parts of the head for about 15 seconds. Then they aligned their bodies in opposite directions with the heads just side by side for penal insertion. The duration of the copulation could not be timed precisely, but the intimate contact lasted 15 to 30 s. When mating ended the bodies shook as if the penis had to be redrawn with force out of the partner. The eggs were in colourless, transparent, gelatinous strings of a length of 9–15 mm and a width of up to 1.7 mm ( Fig. 2 I View Fig ). The strings contained 2000–4000 eggs in a spiral chain ( Fig. 2 J View Fig ). The capsules of fresh eggs were touching each other in the spiral, but not those in neighbouring curves. Fresh, still uncleaved ova yellowish; developing embryos were pale. The egg capsules measured 108.2 ± 3.6 µm by 90.2 ± 6 µm (N = 36), diameters of uncleaved ova 61.1 ± 1.7 µm (N = 22). In captivity, the egg masses were deposited both by submerged and emerged individuals on the mud or on Vaucheria sp. The eggs developed to free-swimming veliger larvae within four days (temperature 28–35°C).
Etymology. – The specific name refers to the striking character that the dorsal side is crowned by the renal shield.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |