
Venetoraptor gassenae gen.et sp. nov., 2023
|
publication ID |
https://doi.org/ 10.1038/s41586-023-06359-z |
|
DOI |
https://doi.org/10.5281/zenodo.8263145 |
|
persistent identifier |
https://treatment.plazi.org/id/03C1A953-5C12-FF9D-BAB0-F9DFECBCFAD6 |
|
treatment provided by |
Felipe |
|
scientific name |
Venetoraptor gassenae gen.et sp. nov. |
| status |
gen. et sp. nov. |
Venetoraptor gassenae gen.et sp. nov.
Etymology. The genus name combines the word raptor,plunderer (Latin) in reference to its raptorial beak and grasping hands and the word Veneto in reference to ‘Vale Vêneto’, a touristic locality within the municipality of São João do Polêsine, Rio Grande do Sul, Brazil. The specific epithet honours Mrs Valserina Maria Bulegon Gassen , one of the main people responsible for the foundation of CAPPA/UFSM.
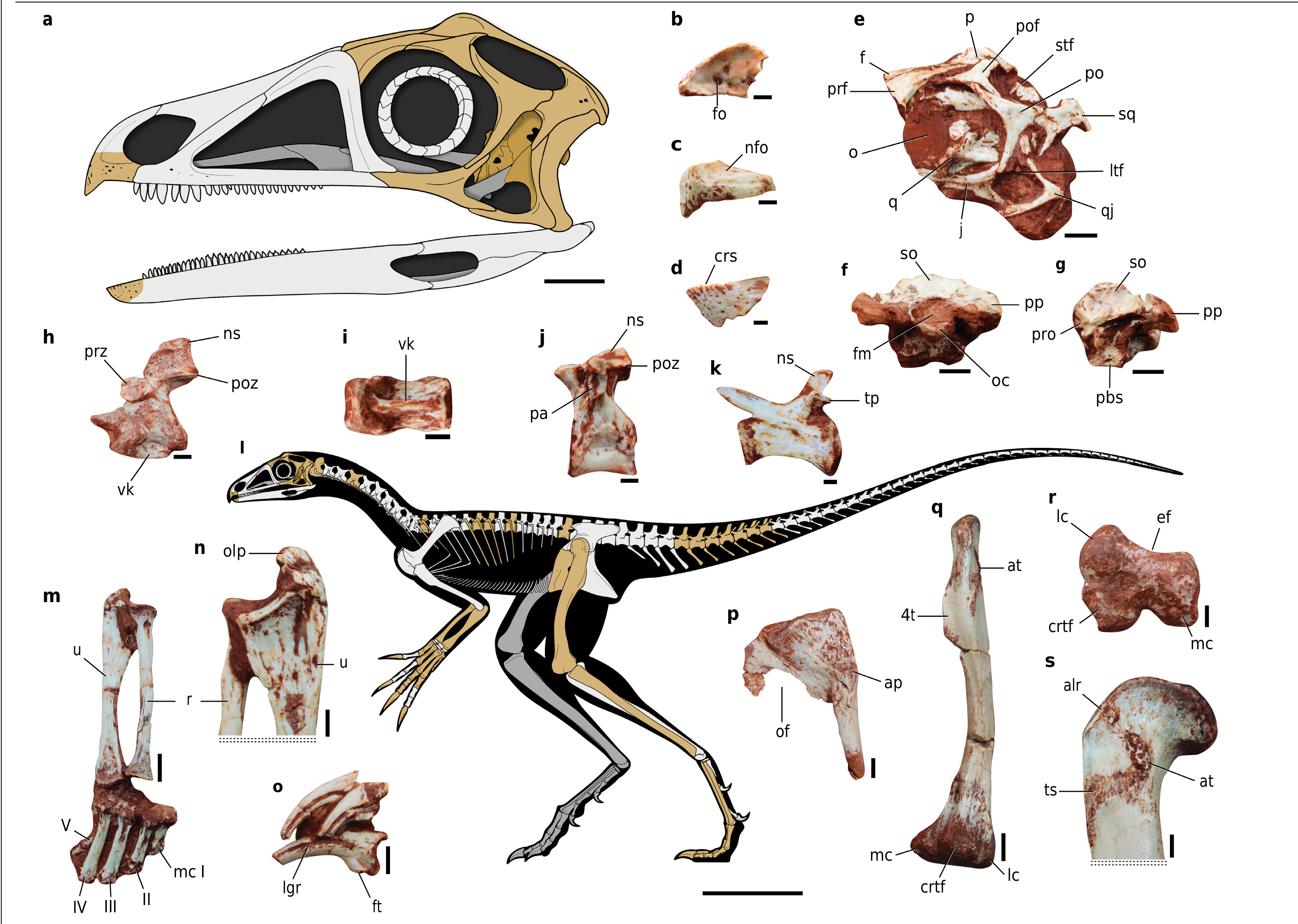
Holotype. CAPPA/ UFSM (Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia da Universidade Federal de Santa Maria) 0356, a partial skeleton of a single individual,including cranial and postcranial remains ( Fig. 1 View Fig ). Locality and horizon. Burial site (29° 39′ 34.2″ S, 53° 25′ 47.4″ W), São João do Polêsine , Rio Grande do Sul, Brazil (Extended Data Fig. 1 View Fig ). The outcrop strata belong to the Santa Maria Formation of the Paraná Basin. The maximum depositional age of biostratigraphically corre- lated beds from a nearby site was recorded as Carnian (233.23 ± 0.73), Ischigualastian reptile age, Late Triassic 13. GoogleMaps
Diagnosis. Differs from other lagerpetids in the following combination of character states (*indicates local autapomorphies):prefrontal contacting postfrontal*;occlusal margin of anterior end of dentary crenulated*;sharply tapering odontoid process of the axis*;dorsal vertebrae with lateral fossa on centrum;metacarpal IVlongerthanmetacarpal III*; pubis with well-developed ambiens process;femur with anterolateral
ridge (≠dorsolateral trochanter) on proximal portion*; trochanteric shelf;ridge for attachment of caudofemoralis muscles (=fourth trochanter); deep extensor fossa on anterior surface of distal portion; concave surface on posterolateral surface of crista tibiofibularis; and fibula with transversely compressed proximal end.
Description
The premaxilla is edentulous (that is, there are no alveoli; Fig. 1b,c View Fig ), with a sharp occlusal margin and a ventrally directed anterior end, resulting in a raptorial beak (Extended Data Fig. 2 View Fig ) resembling that of some birds such as phorusrhacids and falconiforms.The lateral surface of the premaxilla is striated,indicating the potential support of a keratinous covering or rhamphotheca 18, 19 and contrasting with the smooth surface in early pterosaurs 20 and dinosauromorphs 21, 22. A narial fossa occupies most of the base of the internarial process,and four large foramina pierce the palatal surface of the bone.The external naris is not retracted,contrasting with the condition in Triassic pterosaurs 20. The orbit is anteroposteriorly longer than high.The prefrontal is unusually elongated, contacting the parietal posteromedially and the postfrontal posterolaterally,thus excluding the frontal from the external orbital rim ( Fig.1e View Fig ), as probably also occurs in the lagerpetid ‘ Dromomeron ’ gregorii. The triradiate postfrontal is proportionally larger than in most archosauriforms,resembling other lagerpetids 8. This bone forms the posterodorsal margin of the orbit and the anterior margin of the supratemporal fenestra (Extended Data Fig. 3a View Fig ). The braincase ( Fig.1f,g View Fig ) preserves the posterior margin of a deep anterior tympanic recess and the basal tubera are broadly connected to each other medially,as in other lagerpetids 8. The anterior tip of the dentary is edentulous but the occlusal margin is crenulated ( Fig. 1d View Fig ). The lateral surface of the bone adJacent to the occlusal margin is striated and densely pitted ventrally,also indicating a probable keratinous covering.The dentition posterior to the beak, if present,is unknown in Venetoraptor.
As in other lagerpetids the vertebrae are proportionately short along the vertebral column,resembling the condition in early pterosaurs 20. The axis has a well-developed ventral keel( Fig.1h View Fig ).The atlantal centrum and intercentrum are fused to the axis, the former tapering strongly anteriorly. There is a moderately deep fossa on the lateral surface of the dorsal centra ( Fig.1J View Fig ), as in the posterior dorsal vertebrae of Lagerpeton . The preserved dorsal vertebrae have anterior and posterior centrodiapophyseal and postzygodiapophyseal laminae. There is no hyposphene-hypantrum.
The olecranon process of the ulna is robust, enlarged and covered by striations ( Fig. 1n View Fig ). The distal articular surface of the ulna is con- vex whereas the distal end of the radius is concave.The metacarpals increase sequentially in length from digits I to IV ( Fig. 1m View Fig ), as in most pterosaurs and Saltopus 23. Conversely,metacarpal III is the longest in Dromomeron romeri 8 and Scleromochlus taylori 12. Metacarpal V is reduced,slender,positioned ventral to the others and articulates with at least one phalanx (Extended Data Fig. 4a View Extended ). There is a deep fossa on the anterior surface of the proximal portion of the metacarpals and a shallow extensor fossa on their distal ends.The ungual phalanges are proportionally long and trenchant ( Fig. 1o View Fig and Extended Data Fig.3c View Fig ), bearing a large extensor tubercle that is perforated ventrally by a fossa as in D. romeri 8.
The pubis is short and has a distinct dorsolaterally facing anti- trochanteric surface ( Fig. 1p View Fig ), as in Lagerpeton and early pterosaurs (for example, Dimorphodon). The distal tip of the pubis lacks any expansion.The femur ( Fig.1q–s View Fig ) differs from that of other lagerpetids in the presence of an anterolateral vertical ridge on the proximal portion. This ridge is positioned more proximally than the dorsolateral trochanter of other dinosauromorphs 24, reaching the proximal articular surface.There is a rugose anterior trochanter associated with a densely striated trochanteric shelf, as occurs in ‘D.’ gregorii 25. The fourth trochanter is crest-like and symmetrical, contrasting with its absence in Scleromochlus, D. romeri, Dromomeron gigas and early pterosaurs 25, 26. The distal end of the femur has a concave posterolateral surface as in Dromomeron 25. The fibular condyle and crista tibiofibularis form an obtuse angle between each other in distal view, contrasting with the acute angle present in Ixalerpeton , D. romeri and D. gigas 25, 27. The pedal unguals are not strongly recurved and lack a well-developed extensor tubercle. These unguals are slightly larger than those of the manus.By contrast,manual unguals are much larger in pterosaurs 20.
Analyses and discussion
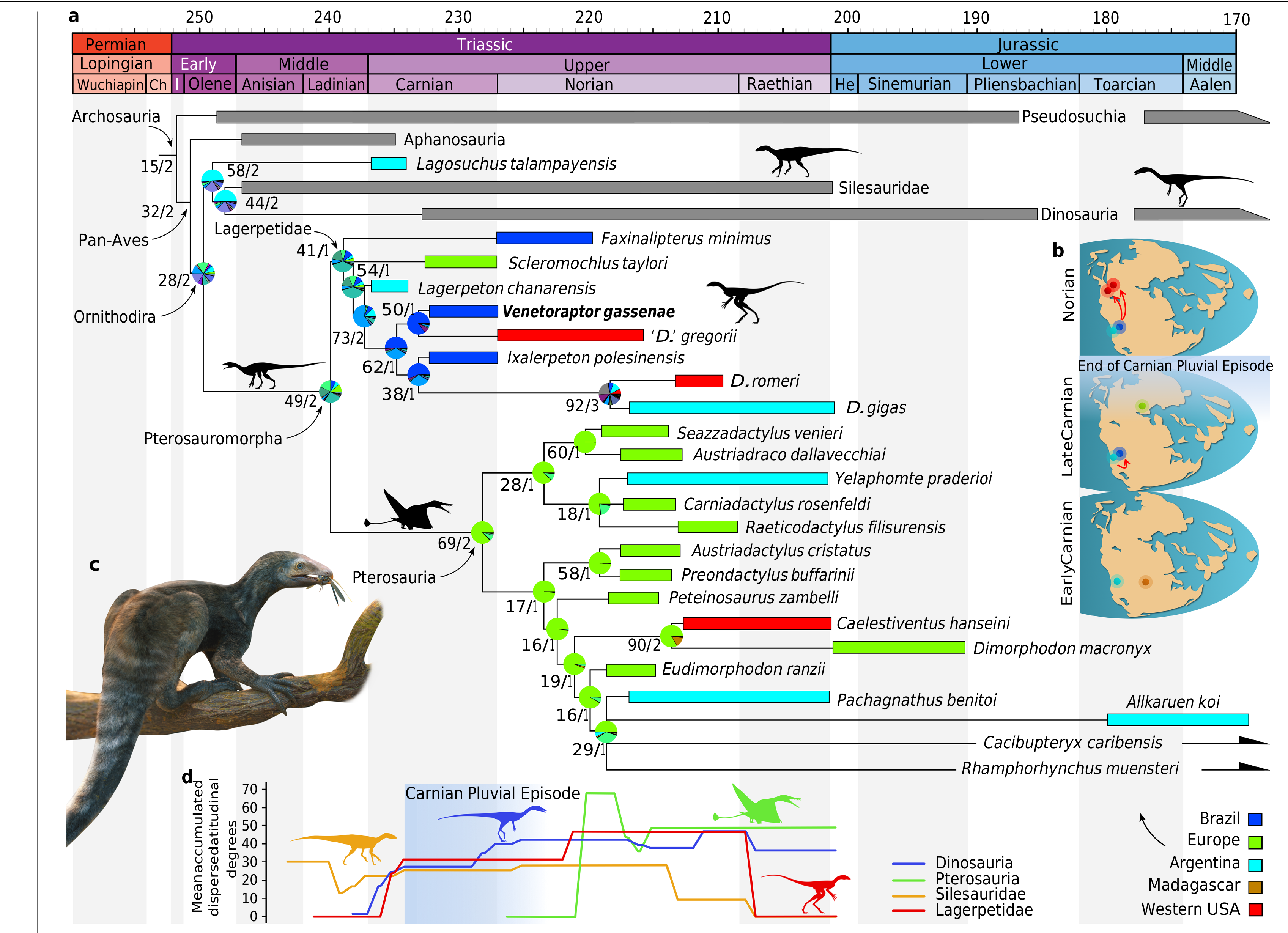
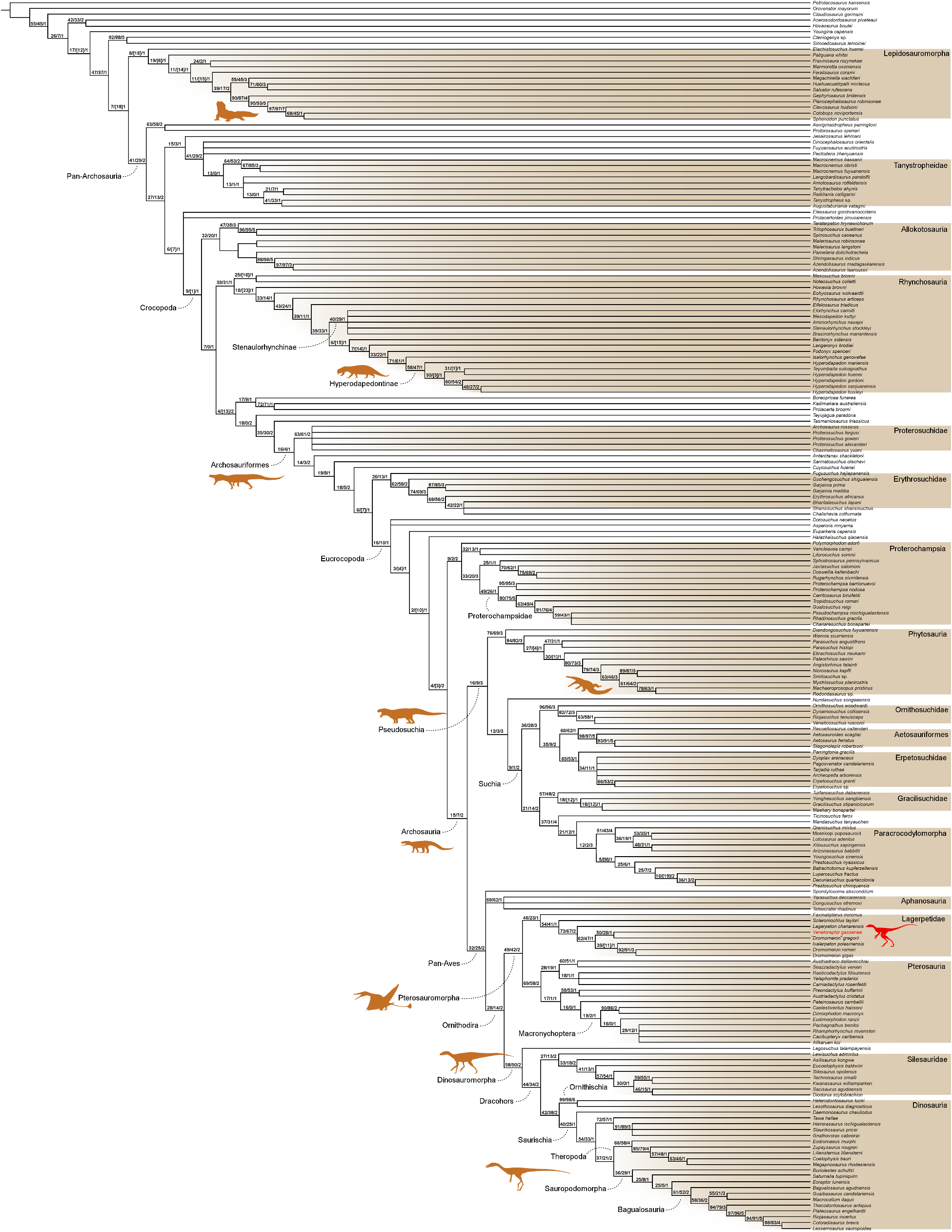
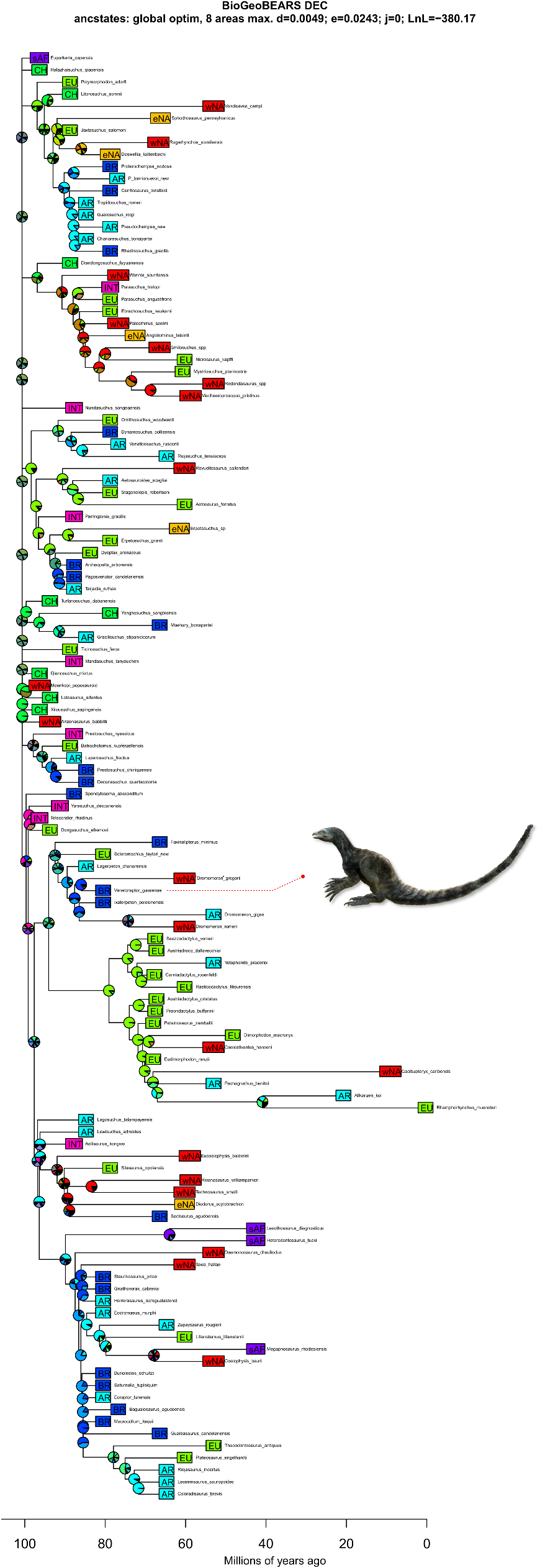
The inclusion of V.gassenae in the most comprehensive phylogenetic analyses of archosauromorph relationships,using both maximum parsimony and Bayesian inference as optimality criteria,nests the new species within Lagerpetidae and lends support to the sister group relationship between Lagerpetidae and Pterosauria 8, 17, 28. Under maximum parsimony, Venetoraptor is the sister taxon to the North American ‘D.’ gregorii in the reduced strict consensus tree ( Fig. 2a View Fig and Extended Data Fig. 5 View Extended ). The coeval Brazilian Ixalerpeton is the sister taxon to a clade comprising D. gigas and D. romeri, from the Norian-Rhaetian of Argentina and North America,respectively.The Bayesian 50% maJority rule tree has a lower resolution within Lagerpetidae,and Venetoraptor is found in a polytomy with all Dromomeron species (Extended Data Fig.6 View Extended ). Ixalerpeton is more shallowly nested than the above-mentioned taxa and in a polytomy that includes the Argentinian Lagerpeton . The occurrence of two North American species deeply nested with South American forms supports a scenario of at least two episodes of dispersion during the Late Triassic ( Fig. 2b View Fig ). Our biogeographical analyses reconstructed both dispersions with a south-to-north direction from a Gondwanan ancestral area ( Fig. 2a View Fig and Extended Data Fig. 7 View Extended ). The apex of dispersed latitudinal degrees for lagerpetids occurred during post-Carnian times, soon after the end of the Carnian Pluvial Event ( Fig.2d View Fig ). This dispersion model closely resembles that of dinosaurs 29, suggesting that the same climatic barriers affected the radiation of both clades.By contrast,silesaurids show a sustained amount of latitudinal dispersion between the Middle-early Late Triassic,and in pterosaurs the peak is delayed until well into the early-middle Norian. Thus,Triassic ornithodirans had a much more complex biogeographic history than previously appreciated.
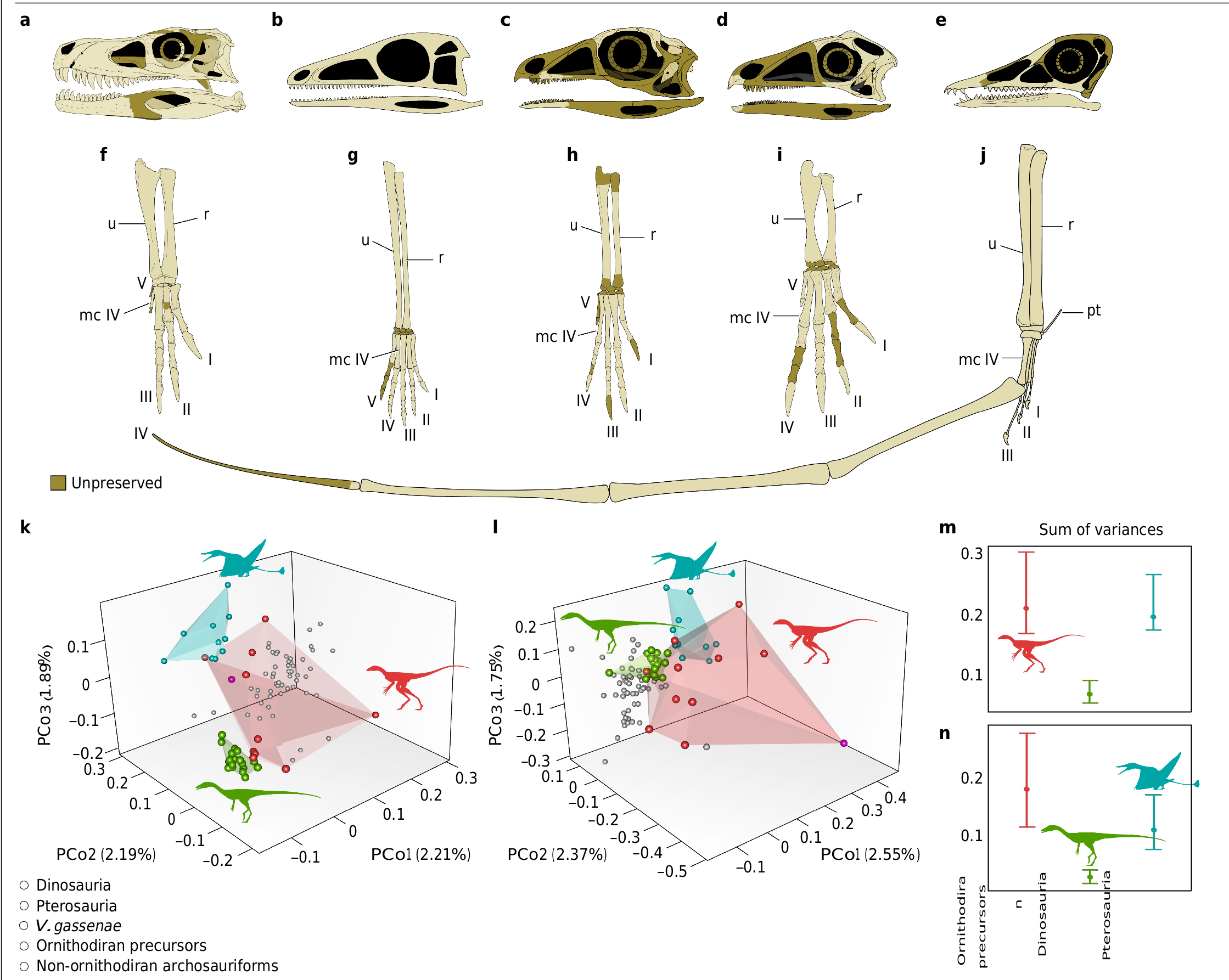
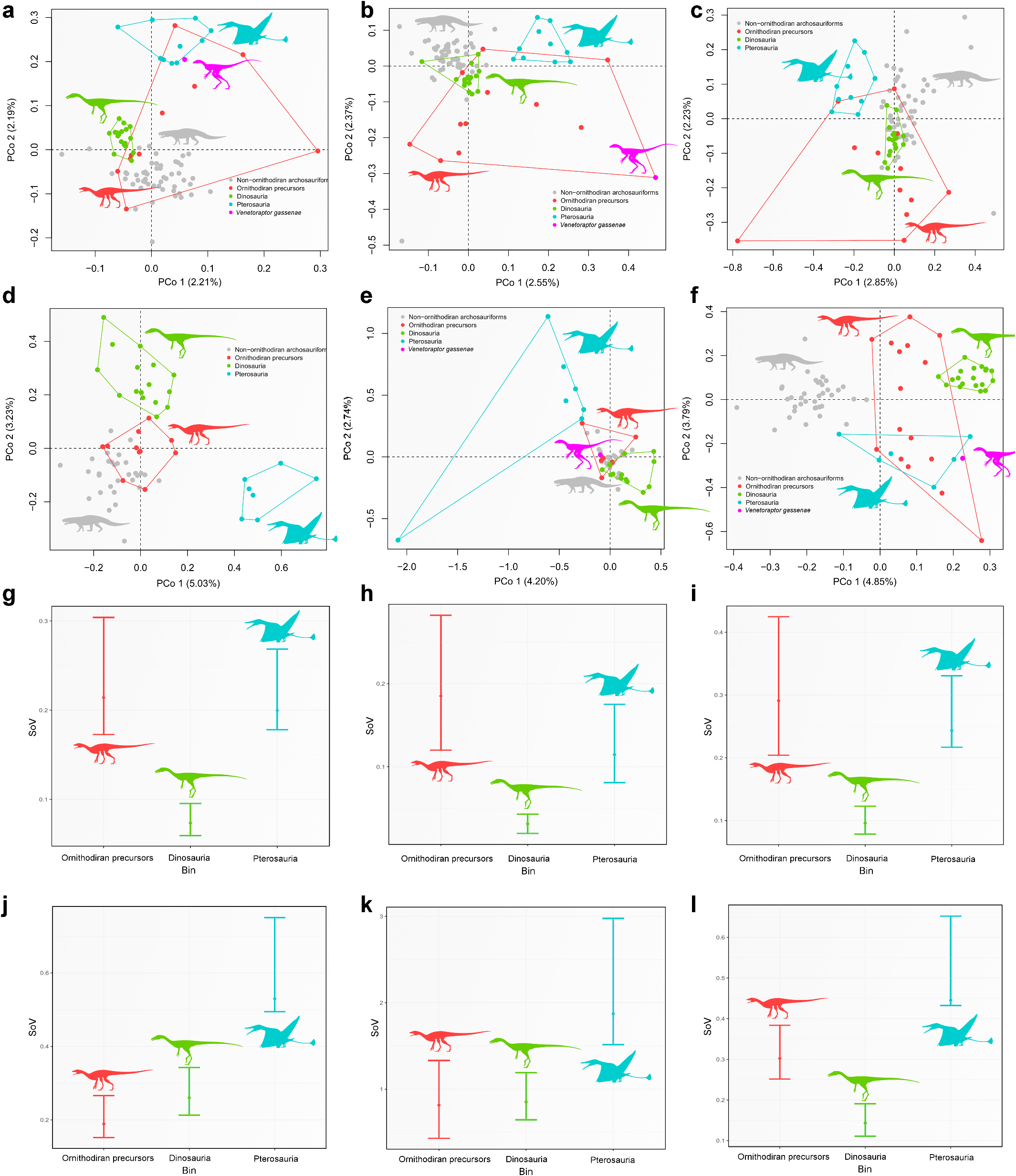
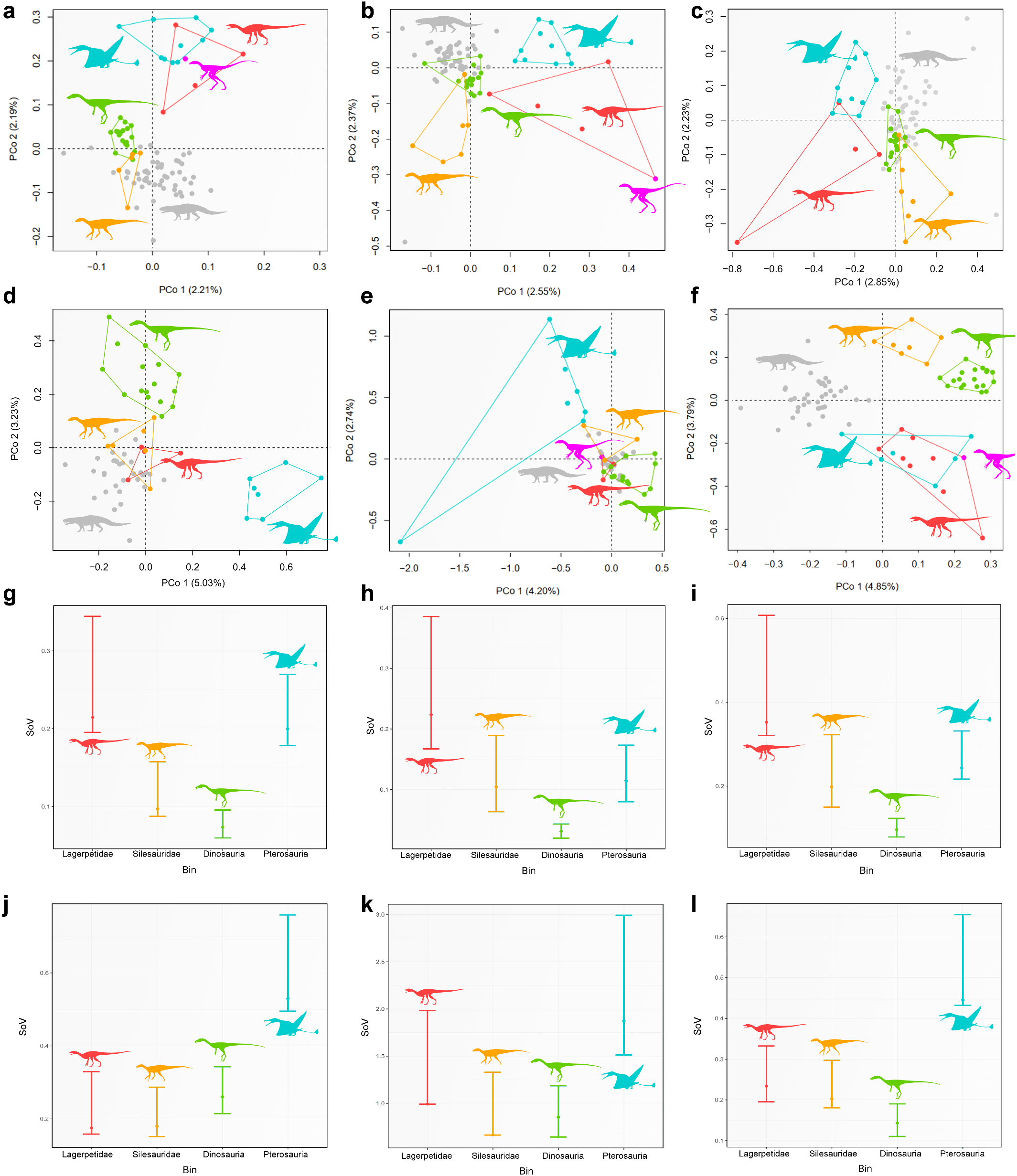
Whereas the phylogenetic relationships of lagerpetids have received considerable attention recently 8, 11, 16, 28,their biology and ecological role remain obscure.The presence of Venetoraptor in the same fossiliferous site that yielded Ixalerpeton represents the first robust evidence of sympatric lagerpetid species.This is particularly interesting because this site, located within the Hyperodapedon Assemblage Zone, also includes one of the oldest dinosaurs(that is,Buriolestes 7) and demon- strates the presence of an already diverse ornithodiran assemblage by the Carnian.Moreover,the new species indicates a previously unappre- ciated ecological diversity among early ornithodirans (=pterosaur and dinosaur precursors, hereafter ‘ornithodiran precursors’),with medium- to small-sized species (femoral length of Venetoraptor is 42 mm versus Ixalerpeton at 69 mm) and different rostral morphologies ( Fig. 3c,d View Fig ). Associated with the discovery of other new species (for example,I.polesinensis 7; Kongonaphon Kely 9) and systematic and morphological re-evaluation of some taxa (for example,I.polesinensis 8; Faxinalipterus minimus 10; Scleromochlus taylori 12), Venetoraptor shows a hidden ecomorphological disparity within Lagerpetidae ( Fig. 3 View Fig and Extended Data Figs.8 View Extended and 9 View Extended ).
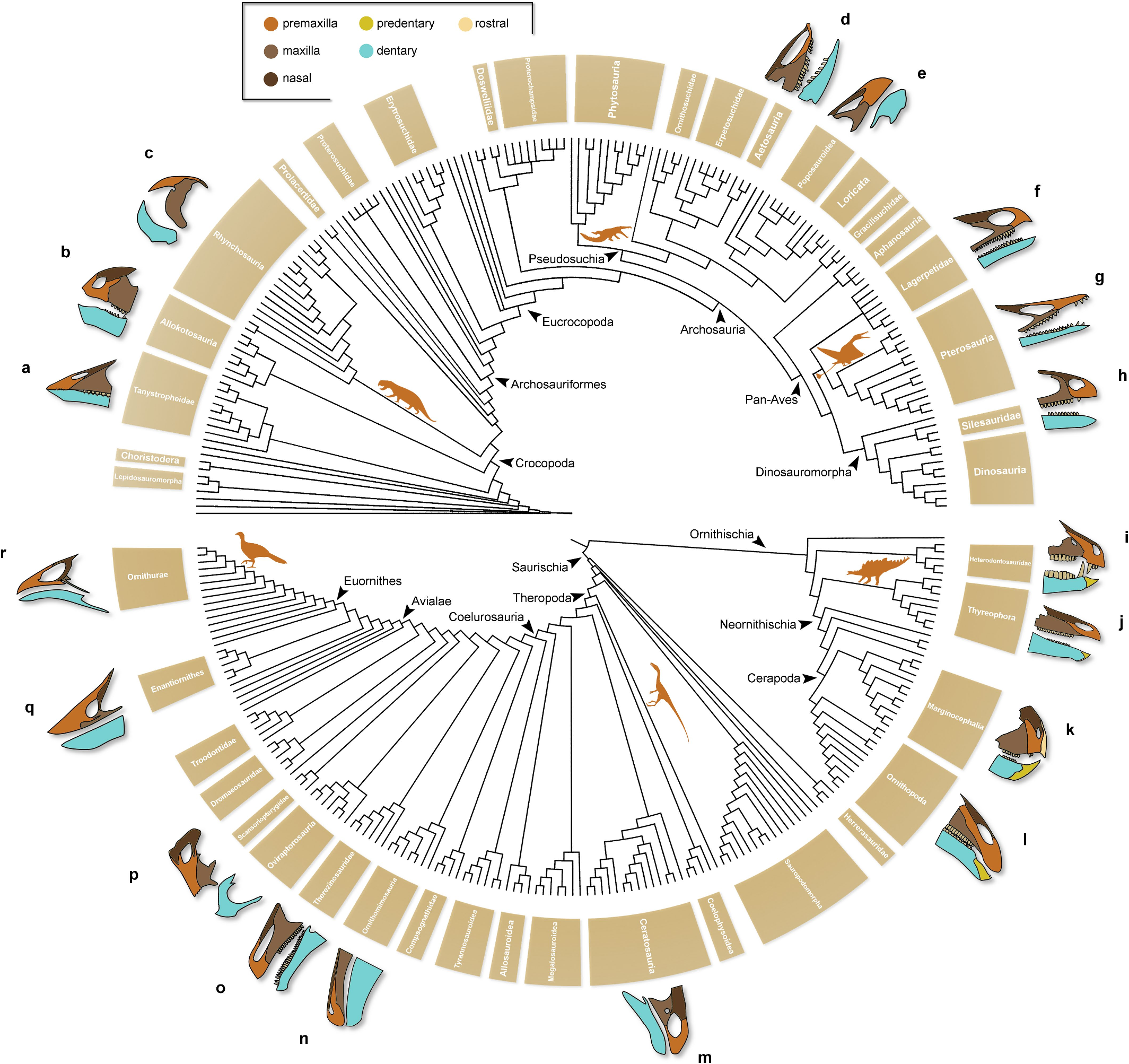
One of the most conspicuous features of Venetoraptor is the presence of a raptorial-like beak.Whereas the main forces that drove edentulism in birds—one of the most studied clades of beaked animals—remain controversial,its functional spectrum is far wider than feeding and foraging 30, 31. This structure plays roles in sexual display, vocalization and thermoregulation,among others 31. The Triassic Period witnessed several episodes of edentulism and origination of beak-like structures within Archosauromorpha 8, 32 – 35 (Extended Data Fig. 10 View Extended ). Neverthe- less, the most numerous episodes of independent beak acquisitions occurred within the avian line of Archosauria.Lagerpetids and silesaurids developed an edentulous and sharp anterior tip of the dentary 8, 36 whereas at least one silesaurid lost part of the premaxillary dentition 22. However, Venetoraptor differs from all other early ornithodirans in the presence of a dorsoventrally taller,ornamented beak with pos- sible support of a rhamphotheca.Furthermore,during the Mesozoic Era distinct lineages of dinosaurs evolved a wide range of beak-like structures 37, 38, including the bird’s beak 39. Pterosaurs evolved distinct kinds of beak-like structures,some forms of which were entirely edentulous during the subsequent Jurassic and Cretaceous Periods 40. The discovery of Venetoraptor expands the morphological spectrum of beaks within Pterosauromorpha,highlights the independent occurrence of this feature in the two main pterosauromorph lineages (that is, Lagerpetidae and Pterosauria ) and shows the first evidence of a raptorial-like beak in this clade. In birds, similar raptorial-like beaks are associated with disparate functions,such as tearing flesh and hard fruit consumption 41. The ecological role and evolutionary advantage of such a beak in Venetoraptor are uncertain.
This new species increases the ecomorphological diversity of the early ornithodiran forelimb.The hands of Venetoraptor are unique and highly specialized,being proportionally large and bearing scythe-like claws ( Fig. 3i View Fig ). Curiously, whereas other lagerpetids have a proportionally shorter metacarpal IV 8, 12 this is the longest metacarpal in V. gassenae. The non-obligatory quadrupedalism is interpreted by us as one of the main forces that drove the evolution of forelimb diversity within Ornithodira during the Late Triassic, which ranged from the non-volant manus with variable number and size of digits to the pterosaur wing.This is clear when the anterior zeugopodium and autopodium morphospace area occupied by Pan-Aves (=Avemetatarsalia) is compared with the considerably smaller area occupied by other archosauriforms (Extended Data Fig.7e View Extended ). Whereas the hindlimb of ornithodirans has been exhaustively investigated,new well-preserved and peculiar forelimbs such as that of Venetoraptor uncover an intriguing field of exploration.Such an enlarged manus bearing sharp claws with deep extensor tubers provides clues to the behaviour of this reptile. Some authors have suggested that the forelimbs of lagerpetids achieved functions other than ground-dwelling locomotion 8, such as climbing and manual processing of food resources.Thus,the recurved beak and grasping hands may have been employed to deal with putative prey and/or scansoriality (that is, the ability to climb).
Quantitative morphological disparity analyses performed here— including the most comprehensive dataset of Triassic Pan-Aves ever assembled—found that the body plan of pterosaurs and dinosaurs evolved as part of a broader morphological diversification of ornithodirans during the Late Triassic.The morphological disparity of the whole skeleton and the skull alone of ornithodiran precursors is signifi- cantly greater than that of Triassic dinosaurs ( Fig.3k–n View Fig ), resembling a previous result based on the whole skeleton 42. This is partially a result of divergent morphologies present between lagerpetids and silesaurids because, when they are analysed as independent groups, their morphological disparity is more similar to that of Triassic dinosaurs (Extended Data Fig. 9 View Extended ). This reflects the wide gamut of feeding appa- ratus that evolved early in the evolutionary history of ornithodirans, ranging from the plesiomorphic ziphodont dentition to specialized beak-like structures 7, 8, 17.The high anatomical variation among Triassic ornithodiran precursors showsthat the flourishing ofTriassic avian-line archosaurs started in early-diverging forms and not after the origin of the volant pterosaurs and cursorial dinosaurs.Venetoraptor and other ornithodiran precursors are part of a previously unexpectedly rich pool of ecomorphological diversity from which were selected two of the most successful vertebrate body plans that ruled the ground and skies during the remainder of the Mesozoic.


| UFSM |
UFSM |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Family |
|
|
Genus |