
Sicyonia laevis Bate, 1881
|
publication ID |
https://doi.org/ 10.5281/zenodo.5394281 |
|
persistent identifier |
https://treatment.plazi.org/id/03C12110-FFEC-FFE9-FCC2-FBB6F202BEEB |
|
treatment provided by |
Marcus |
|
scientific name |
Sicyonia laevis Bate, 1881 |
| status |
|
Sicyonia laevis Bate, 1881 View in CoL
( Figs 65-67 View FIG View FIG View FIG ; 108C, D View FIG )
Sicyonia laevis Bate, 1881: 173 View in CoL ; 1888: 298, pl. 43, fig. 5. — Starobogatov 1972: 412 (clé). — Hayashi 1985a: 143 (clé); 1992: 152 (clé), 161, figs 87b, 88b, 90b.
Eusicyonia laevis – Burkenroad 1934a: 71. — Anderson & Lindner 1945: 318 (liste).
Eusicyonia nebulosa Kubo, 1949: 454 View in CoL , figs 8N, 48C, 77B, H, 79K, 156G, 157.
Sicyonia nebulosa View in CoL – Starobogatov 1972: 412 (clé), pl. 11, fig. 153a, b.
Sicyonia neblosa (sic) – Hayashi 1985c: 260, figs 92b, 93b, 95b.
Non Sicyonia laevis View in CoL – de Man 1911: 118; 1913, pl. 10, fig. 41a, b; 1922: 11, pl. 1, fig. 6, 6a (= S. parvula (de Haan, 1844)) View in CoL .
Non Sicyonia laevis View in CoL – Rathbun 1906: 908, pl. 20, fig. 7 (= S. truncata ( Kubo, 1949)) View in CoL .
MATÉRIEL TYPE. — Bate (1881) mentionne un seul spécimen, un mâle, capturé à la station 219 du Challenger. En fait cet holotype, conservé au NHM, est une femelle.
LOCALITÉ TYPE. — Près des îles Admiralty, au nord de la Papouasie-Nouvelle-Guinée (1°54’S, 146°39’40”E), par 274 m de profondeur.
MATÉRIEL EXAMINÉ. — Madagascar. Vauban, chalutage 5, 12°44,8’S, 48°10,6’E, 570- 563 m, 5.III.1971, 1 10,0 mm (MNHN-Na 13489).
Japon. Tosa Bay , 200-250 m, 3-14.XI.1983, K. Sakai coll., 1 8,8 mm. — Ibidem, 175 m, 3.XII.1997, 1 10,3 mm ( NANSEI). — Ibidem, 200 m, 4.XII.1997, 1 7,4 mm ( NANSEI). — Ibidem, 200 m, 14.XI.1997, 2 6,2 et 7,5 mm ; 2 7,4 et 9,3 mm ( NANSEI). — Au large de Izu Oshima, 34°36’N, 139°26’E, 121-126 m, 17.X.1991, 1 5,8 mm GoogleMaps ; 1 7,7 mm. — Ibidem , 187-204 m, 18.X.1991, 1 6,3 mm. — Central Honshu, Kushimoto, 150 m, 30.XII.1991, S. Nagai coll., K. Nomura pres., 1 6,7 mm. — Ibidem, 100 m, 1.I.1992, S. Nagai coll., K. Nomura pres., 1 7,2 mm. — Ibidem, 250 m, 22.III.1992, S. Nagai coll., K. Nomura pres., 2 5,3 et 9,8 mm. — Ibidem, 200 m, 6.VI.1993, S. Nagai coll., K. Nomura pres., 2 5,0 et 5,4 mm ; 2 5,4 et 6,1 mm. — Ibidem , 180 m, 29.V.1994, S. Nagai coll., K. Nomura pres., 2
5,5 mm et abimé; 1 6,9 mm (MNHN-Na 13490); 9 4,9-7,0 mm et abimée (tous ex SMP- 1015). — Au large de Shionomisaki, 120 m, 8.VIII.1990, 2
4,5 et 5,0 mm; 1 5,5 mm (ex SMP- 269). — Ibidem, 100 m, 20.I.1992, 1 7,0 mm (ex SMP- 528). — Ibidem, 200 m, 6.VI.1993, 2 3,8 et 5,2 mm; 2 4,7 et 4,8 mm (ex SMP- 871). — Ibidem, 160 m, 17.IV.1995, S. Nagai coll., K. Nomura pres., 3 6,7, 6,8 mm et abimée.
Mer de Chine orientale. Yoko-Maru, 30°08,1’N, 127°40,2’E, 192 m, 6.VIII.1996, T. Kosuge coll., 2 5,1 et 5,3 mm ( CBM-ZC 2962).
Philippines. MUSORSTOM 2, stn CP 1, 14°00,3’N, 120°19,3’E, 188-198 m, 20.XI.1980, 1 7,8 mm.
Indonésie. KARUBAR, îles Kai , stn DW 01, 5°46’S, 132°10’E, 156-305 m, 22.X.1991, 1 4,6 mm. — Stn DW 02, 5°47’S, 132°13’E, 209-240 m, 22.X.1991, 5 4,0- 6,5 mm GoogleMaps ; 3 4,7-8,5 mm. — Stn DW 32, 5°47’S, 132°51’E, 170-206 m, 26.X.1991, 1 6,8 mm. — Stn CP 35, 06°08’S, 132°45’E, 390-502 m, 27.X.1991, 1 6,9 mm. — Stn CP 36, 6°05’S, 132°44’E, 210-268 m, 27.X.1991, 1 7,4 mm; 1 9,3 mm.
Papouasie-Nouvelle-Guinée. Challenger, stn 219, 1°54’S, 146°39’40”E, 274 m, 10.III.1875, 1 9,7 mm, holotype ( NHM).
Îles Chesterfield. MUSORSTOM 5, stn 375, 19°52,20’S, 158°29,70’E, 300 m, 20.X.1986, 1 6,1mm.
Nouvelle-Calédonie. BIOCAL, stn DW 64, 24°47,93’S, 168°09,12’E, 250 m, 3.IX.1985, 2
4,9 et 6,0 mm.
MUSORSTOM 4, stn CP 180, 18°56,8’S, 163°17,7’E, 450 m, 18.IX.1985, 1 6,0 mm. — Stn DW 181, 18°57,2’S, 163°22,4’E, 350 m, 18.IX.1985, 1 5,0 mm (MNHN-Na 13491).
CHALCAL 2, stn CP 19, 24°42,85’S, 168°09,73’E, 271 m, 27.X.1986, 1 4,7 mm.
BATHUS 2, stn DW 717, 22°44,02’S, 167°16,58’E, 350-393 m, 11.V.1993, 1 9,1 mm. — Stn CP 737, 23°03,42’S, 166°59,97’E, 350-400 m, 13.V.1993, 2 8,7 et 10,0 mm. — Stn CP 742, 22°33,45’S, 166°25,86’E, 340-470 m, 14.V.1993, 1 8,8 mm.
BATHUS 3, stn CP 847, 23°02,53’S, 166°58,18’E, 405-411 m, 1.XII.1993, 1 9,9 mm.
BATHUS 4, stn CP 906, 19°01,07’S, 163°14,51’E, 339-350 m, 4.VIII.1994, 1 5,7 mm.
HALIPRO 1, stn CP 852, 21°44,98’S, 166°36,82’E, 253-266 m, 19.III.1994, 1 4,6 mm.
SURPRISE, stn 1391, 18°29,8’S, 163°02,8’E, 365 m, 12.V.1999, 2 6,5 et 8,2 mm.
Îles Loyauté. MUSORSTOM 6, stn DW 391, 20°47,35’S, 167°05,70’E, 390 m, 13.II.1989, 2
8,1 et 8,6 mm (MNHN-Na 13582).
Vanuatu. MUSORSTOM 8, stn CP 970, 20°18,56’S, 169°53,19’E, 252-310 m, 21.IX.1994, 1 5,6 mm (MNHN-Na 13581).
Îles Fidji. BORDAU 1, stn CP 1445, 17°10’S, 178°42’W, 350-365 m, 3.III.1999, 2 7,6 mm (MNHN-Na 13544) et 8,2 mm (MNHN-Na 13541). — Stn CP 1446, 17°11’S, 178°42’W, 350-367 m, 3.III.1999, 1 10,0 mm (MNHN-Na 13543). — Stn DW 1469, 19°40’S, 178°10’W, 314-377 m, 8.III.1999, 1 8,6 mm (MNHN-Na 13542).
Îles Wallis. MUSORSTOM 7, stn 610, 13°21,5’S, 176°08,9’E, 286 m, 26.V.1992, 2 5,6 et 6,5 mm.
Îles Tonga. BORDAU 2, stn DW 1583, Vava’u group, 18°37’S, 174°03’W, 327-360 m, 13.VI.2000, 1 4,9 mm. — Stn 1602, centre Ha’apai group, 20°49’S, 174°57’W, 263-320 m, 15.VI.2000, 1 4,2 mm GoogleMaps ; 2 4,8 et 6,1 mm. — Stn DW 1607, seamount, 22°15’S, 175°23’W, 356-367 m, 16.VI.2000, 1 5,1 mm.
DISTRIBUTION. — Madagascar, mer de Chine orientale, Japon, Philippines, Indonésie, Papouasie-Nouvelle- Guinée, îles Chesterfield, Nouvelle-Calédonie, îles Loyauté, Vanuatu, îles Fidji, Wallis et Tonga.
Au Japon, en mer de Chine orientale et aux Philippines, les spécimens que nous avons examinés ont été récoltés entre 100 et 250 m de profondeur (mais Kubo 1949: 456, signale l’espèce au Japon à 350 m environ). En Indonésie on la trouve à ces mêmes profondeurs, à l’exception d’une récolte faite à 390- 502 m. En Nouvelle-Calédonie, aux îles Chesterfield et Loyauté, au Vanuatu et aux îles Fidji, Wallis et Tonga, nous l’avons récoltée entre 250 et 450 m. C’est à Madagascar que l’on relève la plus grande profondeur de récolte (563-570 m).
DESCRIPTION
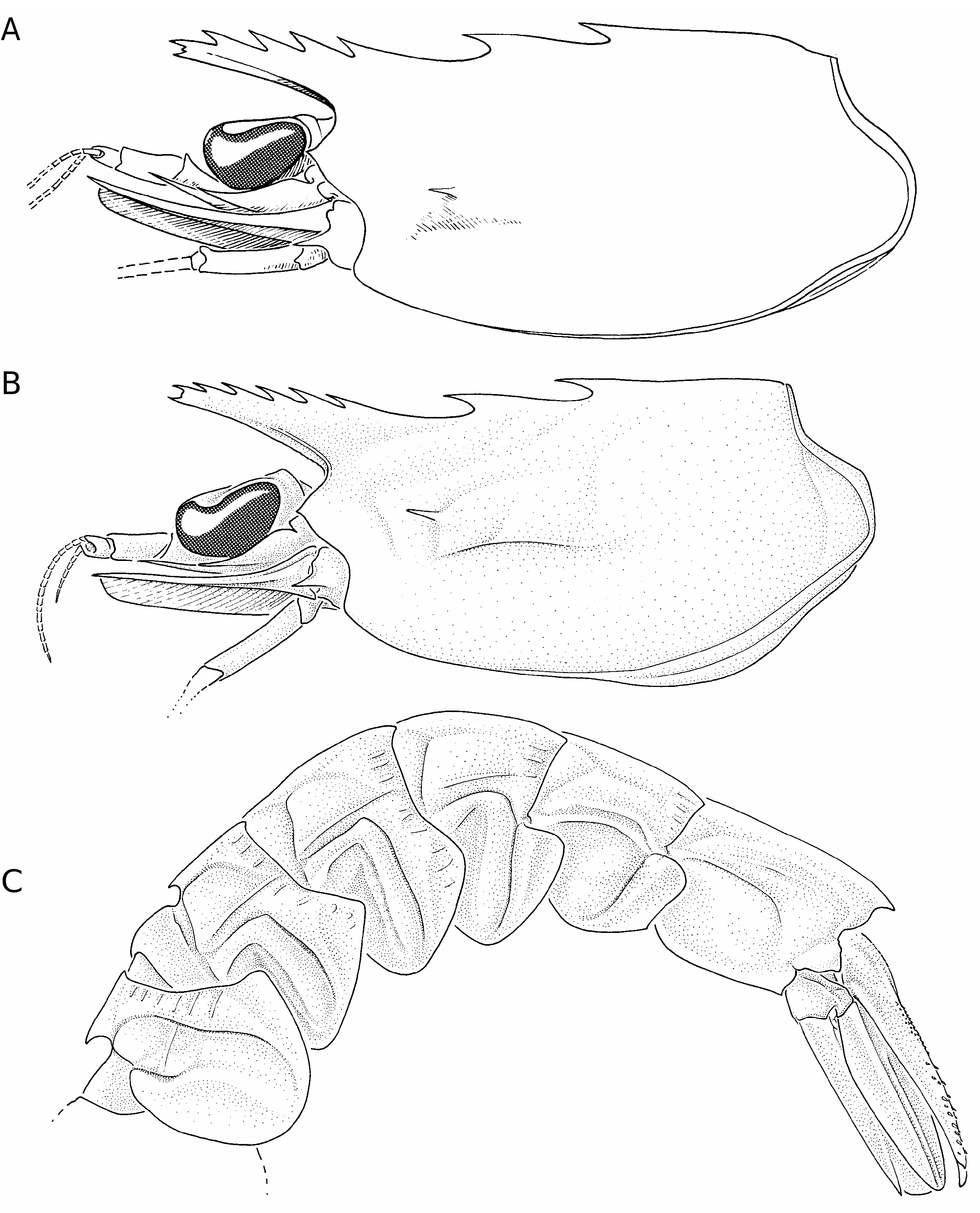
Le rostre légèrement redressé, droit ou très légèrement recourbé ventralement, est plus ou moins grêle suivant les spécimens; son extrémité se situe entre la base et la moitié du deuxième article du pédoncule antennulaire. Le bord dorsal de la carapace porte six ou, plus rarement, sept dents, sans compter la pointe du rostre qui est divisée en deux dents, l’une dorsale et l’autre ventrale et parfois en trois par l’adjonction, entre les deux dents précédentes, d’une dent de taille variable (parfois très petite, parfois aussi grande que celles qui l’encadrent). La première dent se situe à un peu plus des 2/5 de la longueur de la carapace, la troisième juste en arrière de l’orbite; la deuxième est, le plus souvent, un peu plus près de la première que de la troisième (mais elle peut aussi être équidistante ou même très légèrement plus près de la troisième que de la première). Le bord dorsal du rostre porte trois (plus rarement quatre) dents équidistantes entre elles et avec la pointe du rostre.
La carapace porte une épine hépatique fine et assez longue et son lobe infra-orbitaire se termine par un denticule aigu, parfois émoussé ( Fig. 65A, B View FIG ). Les premiers péréiopodes portent une épine subdistale sur le bord interne du basis et de l’ischion. Une épine plus petite s’observe également sur le basis des deuxièmes péréiopodes.
L’abdomen présente, sur le bord dorsal du premier segment, une dent bien développée. Sur le bord dorsal du deuxième segment, on observe une petite dent acérée (parfois émoussée en forme d’angle droit). Le sixième segment porte une dent dorsale terminale. Le bord postéroventral du pleuron du cinquième segment abdominal présente une partie postérieure concave, se terminant du côté ventral par un minuscule denticule, le plus souvent absent. Les sculptures de l’abdomen sont relativement bien marquées: une faible carène longitudinale s’observe au-dessus de la limite entre les tergites et les pleurons des divers segments abdominaux; d’autres petites carènes longitudinales s’observent sur la partie postérieure des segments, la plupart sur les tergites. On observe un sillon postéromédian sur les pleurons des cinq premiers segments et un sillon antéromédian sur les pleurons des trois premiers; celui du premier est à peine marqué. La longueur du sixième segment (mesurée du condyle d’articulation à la pointe du lobe latéral postérieur médian) est, chez les adultes, environ 1,6 fois plus grande que celle du cinquième (mesurée entre les condyles d’articulation).
Le telson se termine en une forte pointe, encadrée par une épine fixe sur chacun de ses côtés, une épine mobile se trouve accolée à cette épine fixe, du côté interne. Les bords ventraux sont inermes mais les limites entre la face dorsale du telson et ses faces latérales sont ornées, sur leurs 3/4 distaux environ, de deux lignes d’une vingtaine d’épines mobiles, irrégulièrement implantées et dont la taille diminue de l’extrémité vers la base du telson.
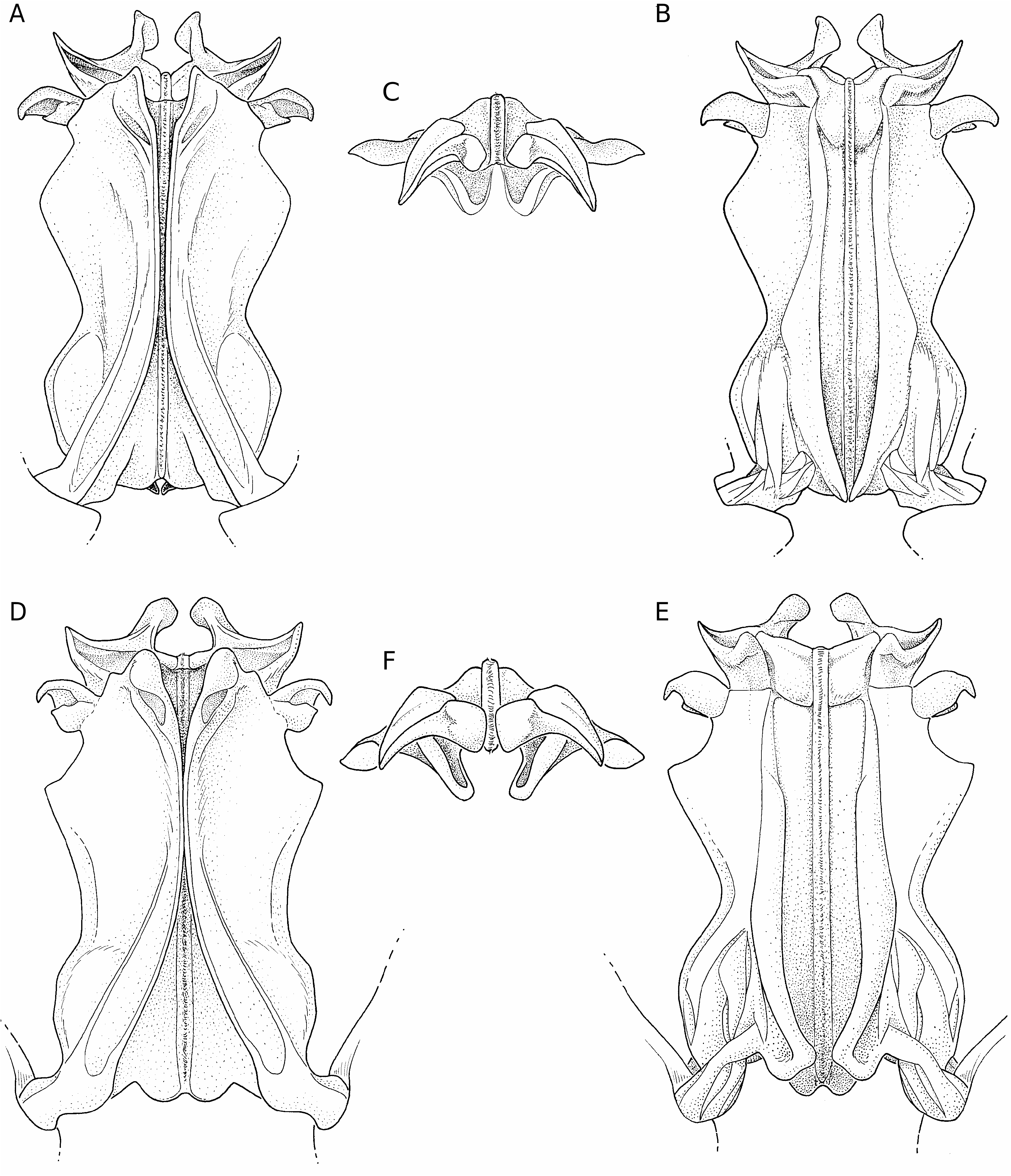
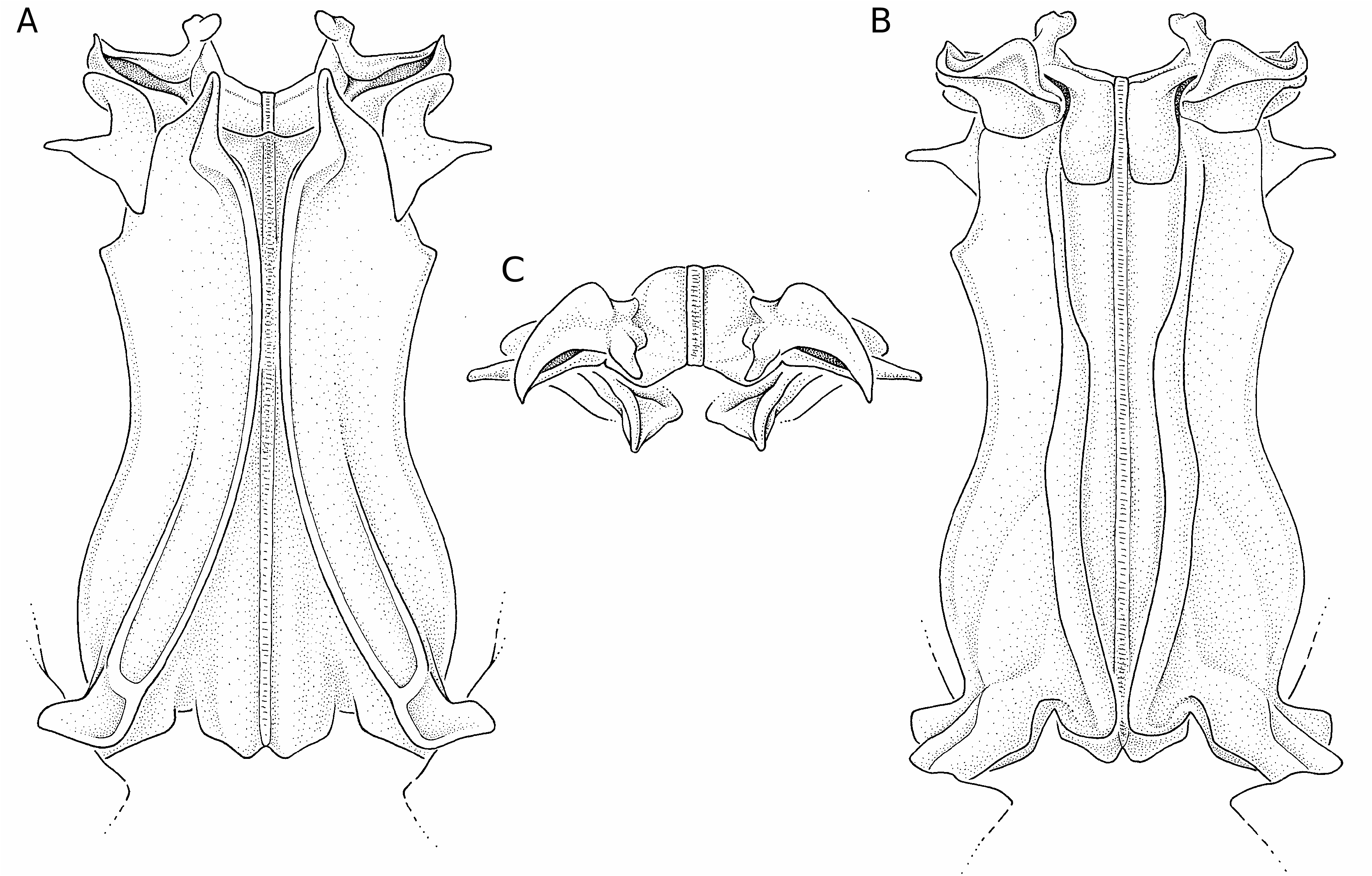
Le thélycum ( Fig. 66 View FIG ) présente une plaque thélycale en forme de très longue pointe, dont les bords latéraux sont fortement sinueux, la plaque étant large dans son tiers basal, puis se rétrécissant rapidement pour, dans son tiers distal, être très fine et acérée. Le rapport L/l de cette plaque semble susceptible de variations importantes, comprises entre 2,5 et 3,0. Cette plaque s’étend jusqu’au niveau des coxas des deuxièmes péréiopodes et est légèrement creusée en cuiller. La plaque postérieure présente deux lobes antérolatéraux dont le bord externe n’est que légèrement convexe; une dépression médiane s’observe dans sa partie antérieure.
Le pétasma ( Fig. 67 View FIG ) se caractérise par le processus distal externe des lobes ventrolatéraux crochu et bifide. Par ailleurs le processus distal interne des lobes dorsolatéraux est bien développé et, antérieurement, dépasse le niveau du processus distal externe; ce dernier est recourbé dorsoventralement. Il n’y a pas de processus latéral en forme de dent aiguë mais les bords latéraux du pétasma présentent un angle très marqué.
Coloration
Elle est très variable. Souvent la partie la plus colorée est la carapace: on observe une large bande transversale brun-rouge qui, sur le bord dorsal, couvre les 2/3 environ de l’espace séparant le bord postérieur de la carapace et la première dent postrostrale puis s’étend jusqu’au bord ventral en laissant l’arrière de la zone branchiale blanche; en avant de cette bande, sur la zone branchiale, on note une bande transversale blanche, pas très large, puis on relève une bande transversale brun-rouge, plus large que la bande blanche, et qui, dorsalement, atteint la base de la deuxième dent postrostrale; la partie antérieure de la carapace est parfois presque uniforme, d’un blanc légèrement teinté de brun, parfois brunrouge avec, de l’arrière à l’avant, une grande bande blanche, suivie d’une bande brune puis d’une nouvelle bande blanche; le rostre est le plus souvent très blanc, sauf dans sa partie distale qui est brun-rouge, mais il peut aussi présenter une suite de bandes brun-rouge et blanches; une bande brun-rouge foncé s’étend sur les pédon- cules antennulaires et les scaphocérites juste audelà de l’oeil. L’abdomen montre souvent des marbrures peu marquées et à dominante blanchâtre sur les segments abdominaux 1-6, tandis que le telson et les uropodes sont entièrement blancs; mais on observe aussi des abdomens très colorés avec quelques taches blanches dans la partie supérieure des pleurons.
Taille
Le plus grand spécimen observé est une femelle dont la carapace mesure 10,0 mm, ce qui correspond à une longueur totale de 36 mm. Le plus grand mâle observé a une carapace de 7,4 mm et une longueur totale de 28 mm.
REMARQUES
Nous avons assez longuement hésité avant de nous décider à mettre S. nebulosa en synonymie avec S. laevis . Il nous avait semblé, au départ, que S. nebulosa pourrait se distinguer par un rostre habituellement plus fort, surtout à sa base, que chez S. laevis et une plaque thélycale plus massive, son rapport L/l n’excédant pas 2,5 contre souvent 3 chez S. laevis . Mais les pétasmas très semblables ( Fig. 67 View FIG ) et, surtout, les variations observées dans chacun des deux groupes nous ont dissuadé de séparer ces espèces. On notera d’ailleurs qu’il ne semble pas que Kubo ait été averti de l’existence de S. laevis lorsqu’il a décrit S. nebulosa car il n’a pas comparé les deux espèces. D’autres considérations sur cette question seront trouvées dans les remarques incluses dans le chapitre relatif à S. rotunda n. sp.
Curieusement de Man, à deux reprises (1911: 118; 1913, pl. 10, fig. 41a, b et 1922: 11, pl. 1, fig. 6, 6a), a identifié à S. laevis des spécimens qui présentent un lobe infra-orbitaire sans épine et un deuxième segment abdominal sans dent dorsale. Le réexamen de ces spécimens a montré que, dans les deux cas, il s’agissait de spécimens de S. parvula .
S. laevis est très proche de S. rocroi n. sp. mais s’en distingue par:
– le lobe infra-orbitaire se terminant en pointe (au lieu d’être régulièrement arrondi);
– le bord dorsal du deuxième segment abdominal portant une petite dent aiguë (au lieu d’une dent en forme d’angle droit à sommet arrondi), précédée d’une encoche assez large (au lieu d’une encoche plus étroite) ( Figs 65C View FIG ; 60B View FIG );
– le processus antérolatéral externe du lobe ventrolatéral du pétasma dont la partie antérieure est nettement plus longue que la partie postérieure, au lieu du contraire ( Figs 67 View FIG ; 62 View FIG ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sicyonia laevis Bate, 1881
| Crosnier, Alain 2003 |
Sicyonia neblosa
| HAYASHI K. - I. 1985: 260 |
Sicyonia nebulosa
| STAROBOGATOV Y. I. 1972: 412 |
Eusicyonia nebulosa
| KUBO I. 1949: 454 |
Eusicyonia laevis
| ANDERSON W. W. & LINDNER M. J. 1945: 318 |
| BURKENROAD M. D. 1934: 71 |
Sicyonia laevis
| MAN J. G. & DE 1911: 118 |
Sicyonia laevis
| RATHBUN M. J. 1906: 908 |
Sicyonia laevis
| HAYASHI K. - I. 1985: 143 |
| STAROBOGATOV Y. I. 1972: 412 |
| BATE C. S. 1888: 298 |
| BATE C. S. 1881: 173 |