
Benedeniella posterocolpa ( Hargis, 1955 ) Yamaguti, 1963
|
publication ID |
https://doi.org/ 10.5281/zenodo.196223 |
|
DOI |
https://doi.org/10.5281/zenodo.6207324 |
|
persistent identifier |
https://treatment.plazi.org/id/03C11D43-D316-FFFD-FF7F-43999F225780 |
|
treatment provided by |
Plazi |
|
scientific name |
Benedeniella posterocolpa ( Hargis, 1955 ) Yamaguti, 1963 |
| status |
|
Benedeniella posterocolpa ( Hargis, 1955) Yamaguti, 1963 View in CoL
( Figs. 3 View FIGURE 3 C–D, 5B, 8–11)
Synonym: Benedenia posterocolpa Hargis, 1955 .
Type host: Rhinoptera bonasus (Mitchill) [syn: R. quadriloba (Le Sueur) ; see Lawler 1981] ( Myliobatidae ), the cownose ray ( Hargis 1955).
Type locality: Tampa Bay, Pinellas County, Florida, Gulf of Mexico, USA ( Hargis 1955).
Other published host and locality records: R. bonasus, Chesapeake Bay , Atlantic coast of USA ( McMahon 1963); R. bonasus , ‘northern Gulf of Mexico’, USA ( Overstreet 1978); R. bonasus, Punta Gorda, Gulf of Mexico, Florida, USA and on captive R. bonasus at Sea World, Florida, USA ( Thoney 1990); R. bonasus, Ocean Springs , Mississippi, Gulf of Mexico, USA ( Kearn & Whittington 1992; present study); R. bonasus, Champotón , Campeche, Gulf of Mexico, Mexico ( Pulido-Flores & Monks 2005); R. bonasus, Punta Gorda, Gulf of Mexico, Florida and Sea World, Florida, USA (present study); R. bonasus held captive at the New York Aquarium, New York City, USA (hosts initially collected in ‘Virginia’ [pers. comm., Dr A.D.M. Dove]) ( Perkins et al. 2009; present study).
Site of infection: Ventral skin ( Hargis 1955; McMahon 1963; Pulido-Flores & Monks 2005; present study); skin ( Thoney 1990).
Infection details: Not determined for any new material examined here.
Material deposited: SAMA AHC 29908 (1 specimen, 1 slide), USNPC No. 103019 (1 specimen, 1 slide) ex R. bonasus (site unknown), Ocean Springs, Mississippi, Gulf of Mexico, USA (original source, RMO); SAMA AHC 29909 (1 slide, 2 specimens), USNPC No. 103020 (1 slide, 7 specimens) ex R. bonasus (site unknown) Punta Gorda, Gulf of Mexico, Florida, USA (original source, DAT); BMNH 2010.4.16.12-13 (2 slides, 2 specimens), SAMA AHC 29910 (1 slide, 1 specimen), USNPC No. 103021 (1 slide, 1 specimen) ex ventral skin surface of R. bonasus, Punta Gorda, Gulf of Mexico, Florida, USA (original source, DAT); BMNH 2010.4.16.14-21 (8 specimens, 8 slides), SAMA AHC 29911 (11 specimens, 11 slides), USNPC No. 103022 – 103031 (10 specimens, 10 slides) ex ventral skin surface of R. bonasus held captive at the New York Aquarium, New York City, USA (hosts initially collected in ‘Virginia’ [pers. comm., Dr A.D.M. Dove]).
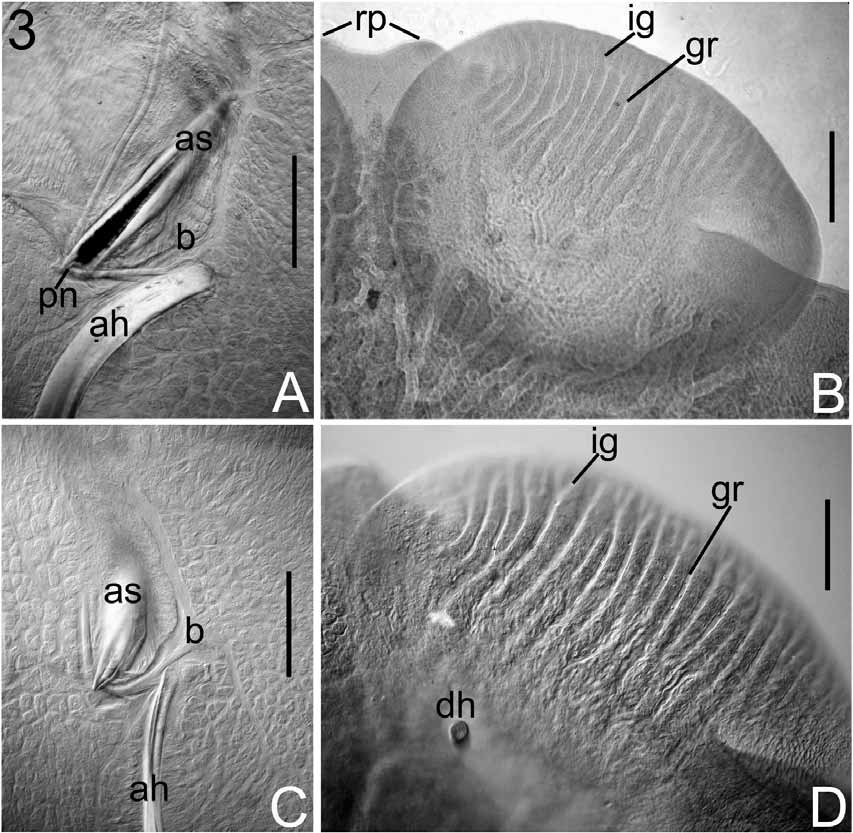
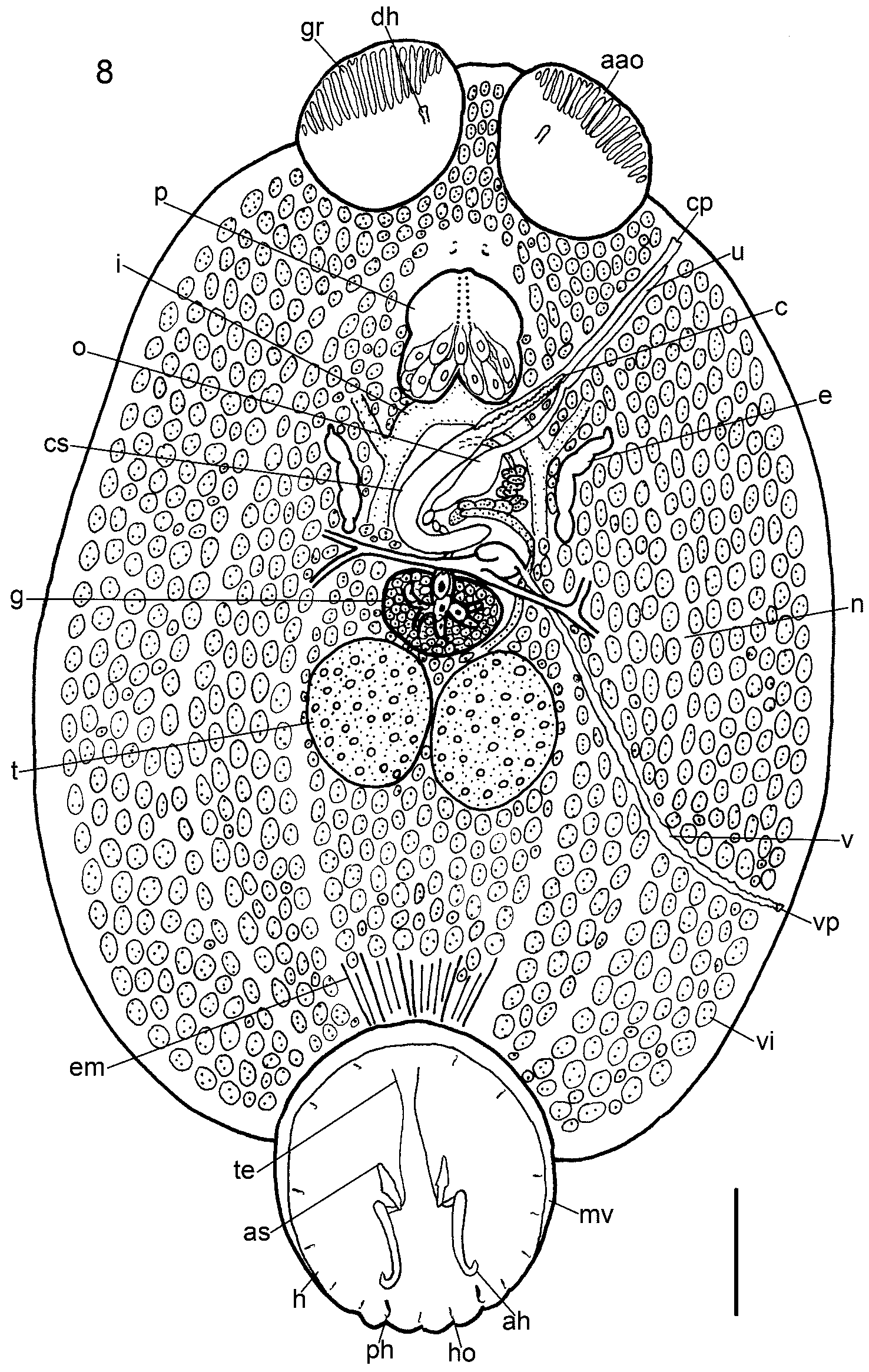
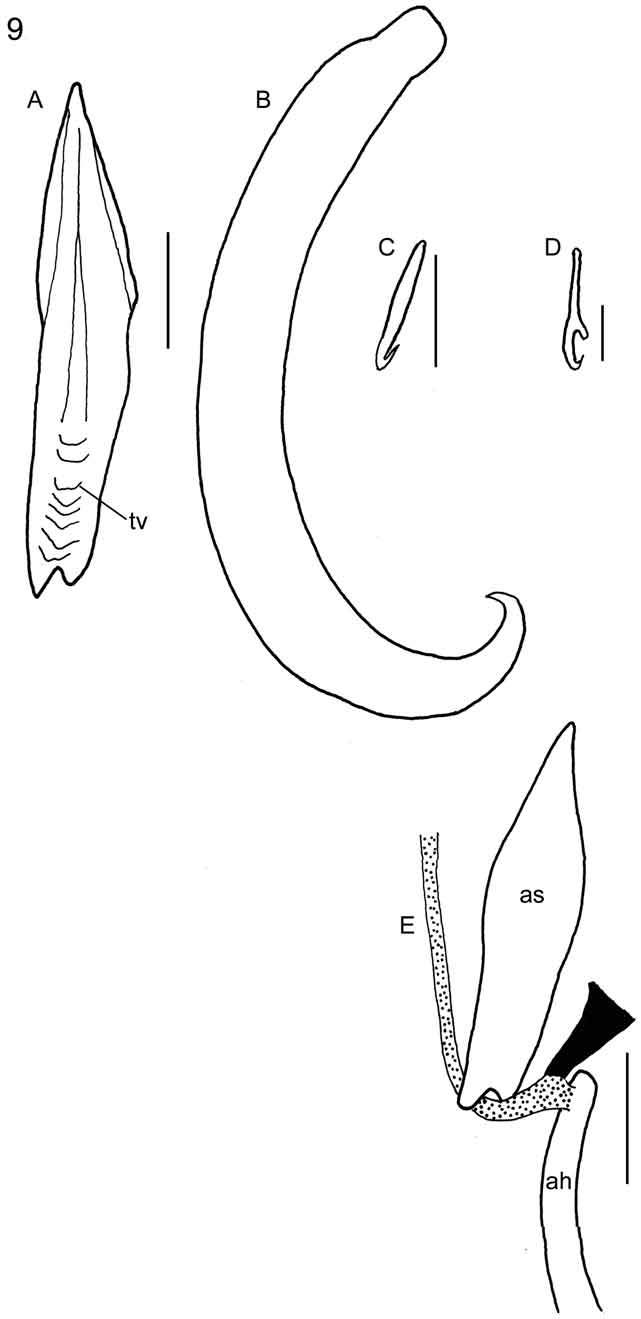
Redescription: Based on 38 flattened wholemounted adult and 6 flattened wholemounted juvenile specimens from various locations off the east coast of the USA. Measurements from 38 flattened wholemounted specimens (all sexually mature), except hooklets measured in flattened wholemounted, mostly juvenile, specimens. Body broadly oval ( Fig. 8 View FIGURE 8 ). Total length (including haptor) and maximum breadth (at a level just posterior to testes): 3.77–17.12 (7.95, n = 38) x 2.44–11.15 (4.54, n = 38) mm. Haptor 1.14–4.19 (2.07, n = 38) x 1.07–3.88 (1.92, n = 38) mm; mostly wider anteriorly, tapering posteriorly ( Fig. 8 View FIGURE 8 ). Accessory sclerites large, 258–863 (378, n = 76) long, near haptor centre; spear head-shaped with transverse ridges proximally ( Fig. 9 View FIGURE 9 A); proximal end with notch ( Figs 3 View FIGURE 3 C, 8, 9A); distal tips sharply-pointed, anteriorly directed ( Fig. 8 View FIGURE 8 ). Anterior hamuli large, stout, strongly recurved distally, 404–1,270 (613, n = 74) long, with hooked distal tips ( Fig. 9 View FIGURE 9 B). Hooked distal tips of one or both anterior hamuli broken off and missing in 21 of 38 mounted specimens leaving blunt tips. Posterior hamuli relatively small, 46–81 (67, n = 74) long, slender with hooked distal tips ( Fig. 9 View FIGURE 9 C); location at haptor periphery indicated by marginal indentation ( Fig. 8 View FIGURE 8 ). Hooklets 13–15 (13, n = 28) long ( Fig. 9 View FIGURE 9 D); positions on posterior and posterolateral borders of haptor indicated by marginal indentations ( Fig. 8 View FIGURE 8 ). Tendons conspicuous, passing posteriorly from extrinsic muscles associated with haptor on each side of body; passing through notch at proximal ends of accessory sclerites before dividing, with part attaching to proximal ends of anterior hamuli and part attaching to ventral haptor tissue ( Figs 3 View FIGURE 3 C, 9E). Marginal valve running anteriorly from position of hooklets IV on each side of haptor extending to anteriormost extremity of haptor; wider anteriorly; absence at posteriormost part of haptor (e.g. Fig. 8 View FIGURE 8 ) from narrowness and damage. Ventral haptor surface apapillate. Pair of elongate, conical structures dorsal to haptor near peduncle observed in stained wholemounts when viewed with dorsal surface uppermost (e.g. Fig. 5 View FIGURE 5 B).
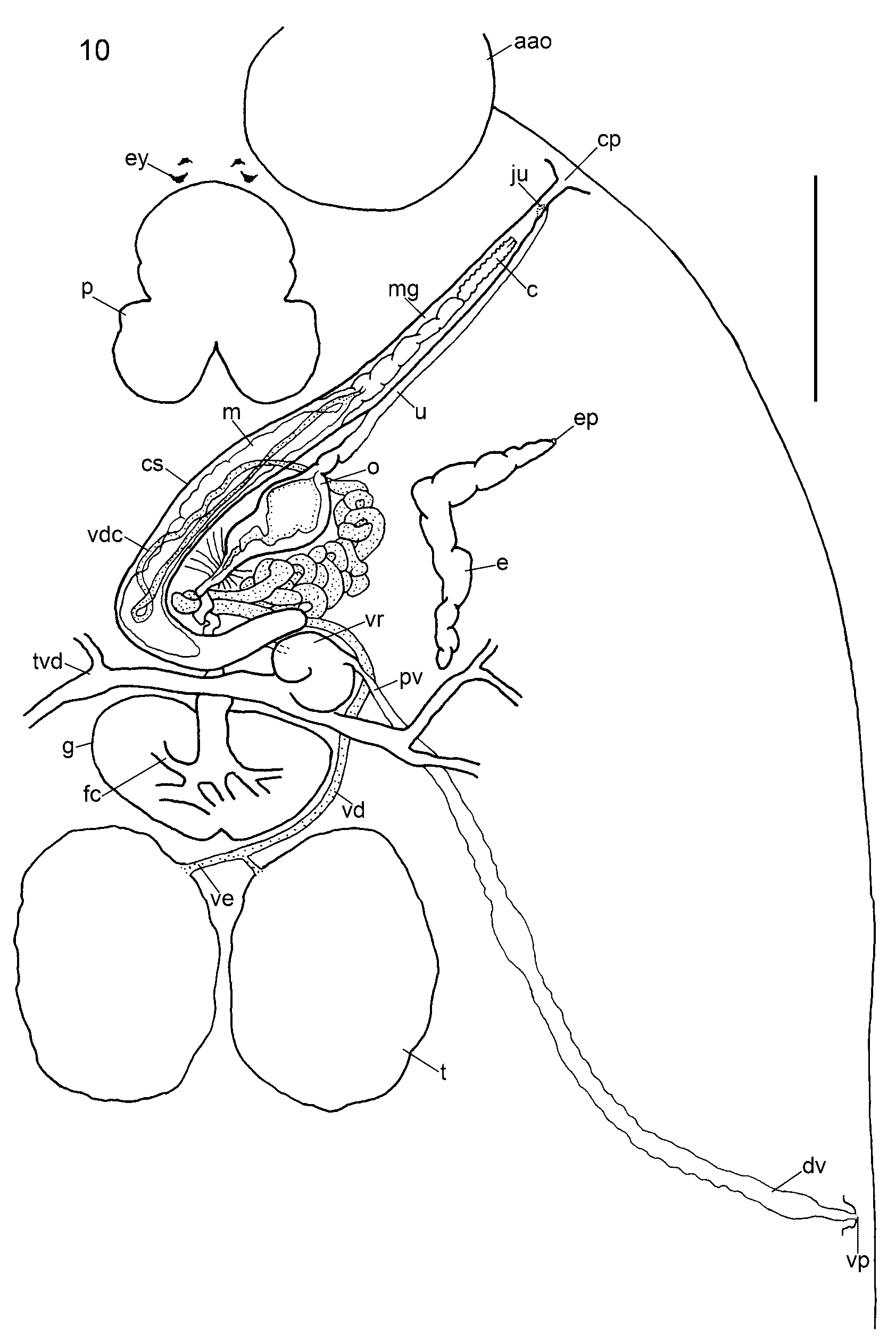
Anterior attachment organs ventral, almost circular, saucer-like discs, 563–2,307 (1,138, n = 64) x 559–2,264 (1,076, n = 64); ventral anterolateral region bearing conspicuous, complete grooves ( Figs 3 View FIGURE 3 D, 8) of variable number, 13–26 (20, n = 34). Number of grooves in juvenile individuals similar to adults. In addition to complete grooves, 0–7 (3, n = 24) incomplete grooves present, of different lengths, occupying positions between complete grooves ( Fig. 3 View FIGURE 3 D). Pair of tapering, horn-like papillae, one horn dorsal to each anterior attachment organ near junction with body ( Figs 3 View FIGURE 3 D, 8); affinity for histological stains (e.g. carmine) in some stained specimens (e.g. Fig. 3 View FIGURE 3 D). Eyes two pairs, pigment-shielded, dorsal, immediately anterior to pharynx. Pharynx conspicuous, 465–2,012 (914, n = 36) x 551–1,796 (852, n = 36); intrinsic gland cells numerous posteriorly. Intestinal caeca branching extensively in body proper, coextensive with and mostly obscured by vitellarium; unclear whether gut caeca confluent posteriorly. Excretory bladders conspicuous, often with lateral bend anteriorly, orientated obliquely towards body margins, each opening dorsally on distinct excretory papilla at level of cirrus base ( Figs 5 View FIGURE 5 B, 8, 10), with some affinity for histological stains.
Testes ovoid, juxtaposed, 452–2,191 (992, n = 74) x 434–1,777 (812, n = 74); penetrated by numerous dorsoventral muscle bundles ( Fig. 8 View FIGURE 8 ). Glands of Goto absent in juvenile and adult specimens. Vasa efferentia short uniting immediately posterior to germarium ( Fig. 10 View FIGURE 10 ). Vas deferens skirting left of germarium, passing dorsal to transverse vitelline duct and proximal part of vagina before numerous intense convolutions occupying bulk of region immediately posterior and left of ootype chamber ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ); entering muscular wall of cirrus sac dorsally at level of anterior region of ootype chamber. Within cirrus sac, vas deferens narrowing, following posterior course dorsally to region of median turn of sac, then returning ventrally to enter base of cirrus ( Fig. 10 View FIGURE 10 ). Widening of vas deferens or presence of sperm-filled reservoir inside cirrus sac not observed. Male accessory gland reservoir single, located ventrally in cirrus sac, not occupying posteriormost region ( Fig. 10 View FIGURE 10 ); source of male accessory secretion not identified; no ducts observed penetrating cirrus sac proximally. Cirrus sac with muscular walls (not shown); distal two-thirds lying diagonally in body posterior to pharynx; proximal third curving medianly, immediately anterior to germarium and vitelline reservoir ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ). Relationship between total length and extent of curvature of cirrus sac with total parasite length observed for B. macrocolpa not apparent for B. posterocolpa . Distal duct from male accessory gland reservoir and distal region of vas deferens uniting near base of cirrus at level of excretory papillae ( Fig. 10 View FIGURE 10 ). Cirrus long, sinuous, sometimes twisted, running anterolaterally inside terminal male genital duct, ventral to separate uterus, opening submarginally, dorsally, on left side of body at level of anterior region of pharynx via common genital pore ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ); common genital papilla not observed. Cirrus eversible outside animal to length of 0.4–2.9 (2.0, n = 30 measured along curves from wholemounted adult specimens) mm.
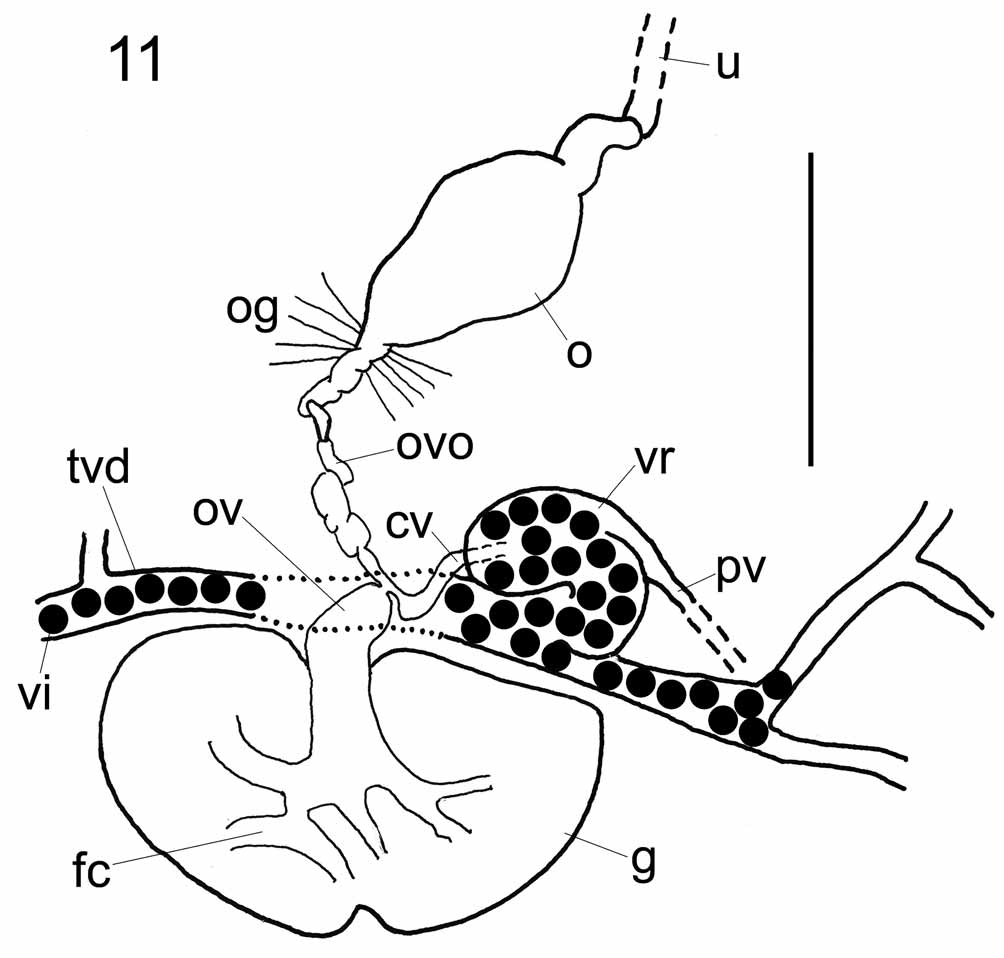
Germarium immediately pretesticular with conspicuous, branched, expanded fertilisation chamber ventrally at mid-line ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ), containing ripe oocytes. Oviduct short, wide, joining short common vitelline duct from vitelline reservoir to form ovo-vitelline duct immediately pregermarial, dorsal to transverse vitelline duct ( Fig. 11 View FIGURE 11 ). Ovo-vitelline duct long, winding, travelling anteriorly to join ootype ( Figs 10 View FIGURE 10 , 11 View FIGURE 11 ). Numerous fine gland ducts entering ovo-vitelline duct at base of proximal region of ootype ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 , 11 View FIGURE 11 ); many small gland cells (not shown) located in curve of cirrus sac, right of ovo-vitelline duct. Ootype with convoluted proximal duct leading to tetrahedral-shaped chamber at level of excretory bladders, joining uterus ( Fig. 10 View FIGURE 10 ). Uterus long, following similar but dorsal course parallel to terminal male genital duct, uniting short distance from common genital pore ( Fig. 10 View FIGURE 10 ; see also Kearn & Whittington 1992). Common genital duct short, opening dorsomarginally at common genital pore at level of anterior part of pharynx ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ). No common genital papilla observed.
Vitelline reservoir ventral, curving anteromedianly near proximal end of cirrus sac. Vagina 1.2–5.7 (2.6, n = 38, measured along curves) mm long; originating proximally via narrow duct from left dorsal margin of vitelline reservoir. No seminal receptacles or proximal specialisations to vagina. Vagina following posterolateral path dorsal to transverse vitelline ducts to a level posterior to testes, opening posteriorly, dorsomarginally ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ). Vitellarium follicular, extending from between anterior attachment organs almost to posterior extremity of body proper, coextensive with intestinal caeca ( Fig. 8 View FIGURE 8 ). Free eggs not observed; in some mounted specimens, eggs observed inside ootype, with tetrahedral shape and appendage; no egg measurements made due to distortion during specimen processing.
Remarks: The anatomy of B. posterocolpa presented here is consistent in most respects with the original description by Hargis (1955) and additional details provided by McMahon (1963). The principal differences reported here relate to the arrangement of the terminal female and male genital systems. Hargis (1955) described separate female, male and vaginal pores in B. posterocolpa whereas in the present study, the same arrangement as described for B. macrocolpa (see above) is confirmed and agrees with the opinion of Kearn & Whittington (1992): in B. posterocolpa , the uterus is very long, running dorsal to, and parallel with, the separate long terminal male genital duct, with the female and male systems uniting distally near the dorsal common genital pore. Hargis’s (1955) description of the uterine pore opening dorsal to the left bifurcation of the major gut branches (see his Plate V, figure 38) was clearly an error. Hargis (1955) also referred to ‘prostate glands’ (= male accessory glands) in 2 large fields close to the excretory bladders, but these glands were not observed in the present study and may be associated with differences in specimen preparation.
The route of conspicuous tendons in the haptor and, after dividing, their partial association with the proximal ends of the accessory sclerites and anterior hamuli and the presence of complete and some incomplete grooves on the anterior attachment organs, were also noted in this redescription of B. posterocolpa . Hargis (1955) illustrated the route of tendons in the haptor of B. posterocolpa (see his Plate V, figure 38), but did not describe them. However Hargis did refer to the anterior attachment organs as follows: ‘anterior half curiously striated’. In the present study, 13–26 complete and 0–7 incomplete grooves were observed.
The presence of horn-like dorsal structures identified on the anterior attachment organs by McMahon (1963) is confirmed here. Other dorsal features (papillae associated with the excretory bladders; elongate conical structures associated with the haptor near the peduncle) are reported here for the first time in B. posterocolpa (e.g. Figs 3 View FIGURE 3 D, 5B, 8). These dorsal features are shared with B. macrocolpa (see above).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |