
Adriaphaenops Noesske, 1928
|
publication ID |
https://doi.org/ 10.5281/zenodo.200269 |
|
publication LSID |
lsid:zoobank.org:pub:20C47092-7D67-4E65-ADE0-A73ED91DBB18 |
|
DOI |
https://doi.org/10.5281/zenodo.6085107 |
|
persistent identifier |
https://treatment.plazi.org/id/03C08791-FFA5-FF92-1499-0C38FAFFFAC0 |
|
treatment provided by |
Plazi |
|
scientific name |
Adriaphaenops Noesske, 1928 |
| status |
|
Genus Adriaphaenops Noesske, 1928 View in CoL
Adriaphaenops Noesske, 1928:5 View in CoL , type species: Trechus antroherponomimus Noesske, 1928 View in CoL by monotypy, type locality: Čatol jama des Bjelasica-Gebirges (weitere Umgebung von Gacko) im nord-ostherzegowinischen Karstlande.
Aphaenopsis View in CoL (sg. Adriaphaenops View in CoL ) Jeannel, 1928:793, Pretner, 1959:79, Casale & Laneyrie, 1982:159, Vigna Taglianti & Sciaky, 1988:166, Pavićević, 1990:365.
Aphaenops View in CoL (sg. Adriaphaenops View in CoL ) Scheibel, 1935:34, Pavićević, 2001:33.
Adriaphaenops Sciaky & Vigna Taglianti, 1990:171 View in CoL , Monguzzi, 1993:238, Casale & Guéorguiev, 1994:421. Drovenik & Peks, 1994:43, Moravec et al., 2003:289, Quéinnec, 2008:157, Quéinnec & Pavićević, 2008:144.
Quéinnec, Pavićević & Ollivier, 2008:154, Lakota et al., 2010:100, Lohaj & Lakota, 2010:78.
Diagnosis. Small to medium-sized trechine genus, L: 3.5–5.75 mm, TL: 3.4–5.35 mm, with aphaenopsoid characters: elongate head and pronotum, elytra ovoid with base strongly narrowed, distinctly wider than head and pronotum, body apterous, depigmented, strongly flattened, more or less pubescent. Antennae and legs elongate, densely pubescent.
Head varying in shape, from almost rounded ( pretneri , perreaui , jasminkoi sp. nov.) to more or less elongate (other species), as long as wide ( zupcense ) or distinctly longer than wide (other species), as wide as pronotum, or slightly narrower. Eyes totally absent. Frontal furrows moderately impressed, incomplete, usually shorter than half of head length, or reaching middle of head ( mlejneki sp. nov.). Usually two pairs of supraorbital setae present, in albanicus sp. nov. and mlejneki sp. nov. anterior pair is absent, in staudacheri , perreaui and njegosiensis sp. nov. both pairs are missing or reduced (indistinguishable from pubescence). Mandibles very long and slender, at apex pointed, right mandible without tooth. Clypeus with four ( antroherponomimus , pretneri , stirni and mlejneki sp. nov.) or three (other species) pairs of setae, labrum with three pairs. Mentum imperfectly fused with submentum. Antennae long and slender, covered with dense pubescence, scape as long as pedicel, antennomeres 3–7 and 8 –11 nearly equally long.
Pronotum elongate, distinctly longer than wide, variously shaped. Basal part as wide as anterior ( pretneri ) or narrower (other species), anterior angles distinct, at apex pointed, or more or less rounded, posterior angles acute, sharp ( zupcense ), or obtuse, more or less rounded (other species). One pair of antero-lateral setae present; basolateral setae absent.
Elytra subovate elongate, with maximum width at middle or apical third, distinctly wider than head and pronotum. Basal part strongly narrowed, pedunculate, shoulders totally effaced. Scutellum small, single pair of basal scutellar setiferous pores present. Elytral striae absent. Pubescence of elytra sparse ( kevser , rumijaensis sp. nov.) or more or less dense (other species), site of stria 3 with four (three discal and one preapical) ( kevser , rumijaensis sp. nov.) or three (two discal and one preapical) dorsal pairs of setae (other species). Humeral group of umbilicate pores not aggregated, first pore of humeral group isolated and situated before level of first discal setae.
Legs long and slender, pro- and mesotarsi short, claws very long and acute, without denticulation, densely pubescent. First two tarsomeres of male protarsi dilated at internal margins.
Aedeagus ( Figs. 23–30 View FIGURES 23 – 30 ) from short and wide ( perreaui , antroherponomimus ) to elongate and slender in lateral aspect, apex variously shaped. Endophallus generally without distinct sclerotised copulatory piece or this piece is only very slightly defined. Parameres from short and wide to long, each with two long and thick setae at apex.
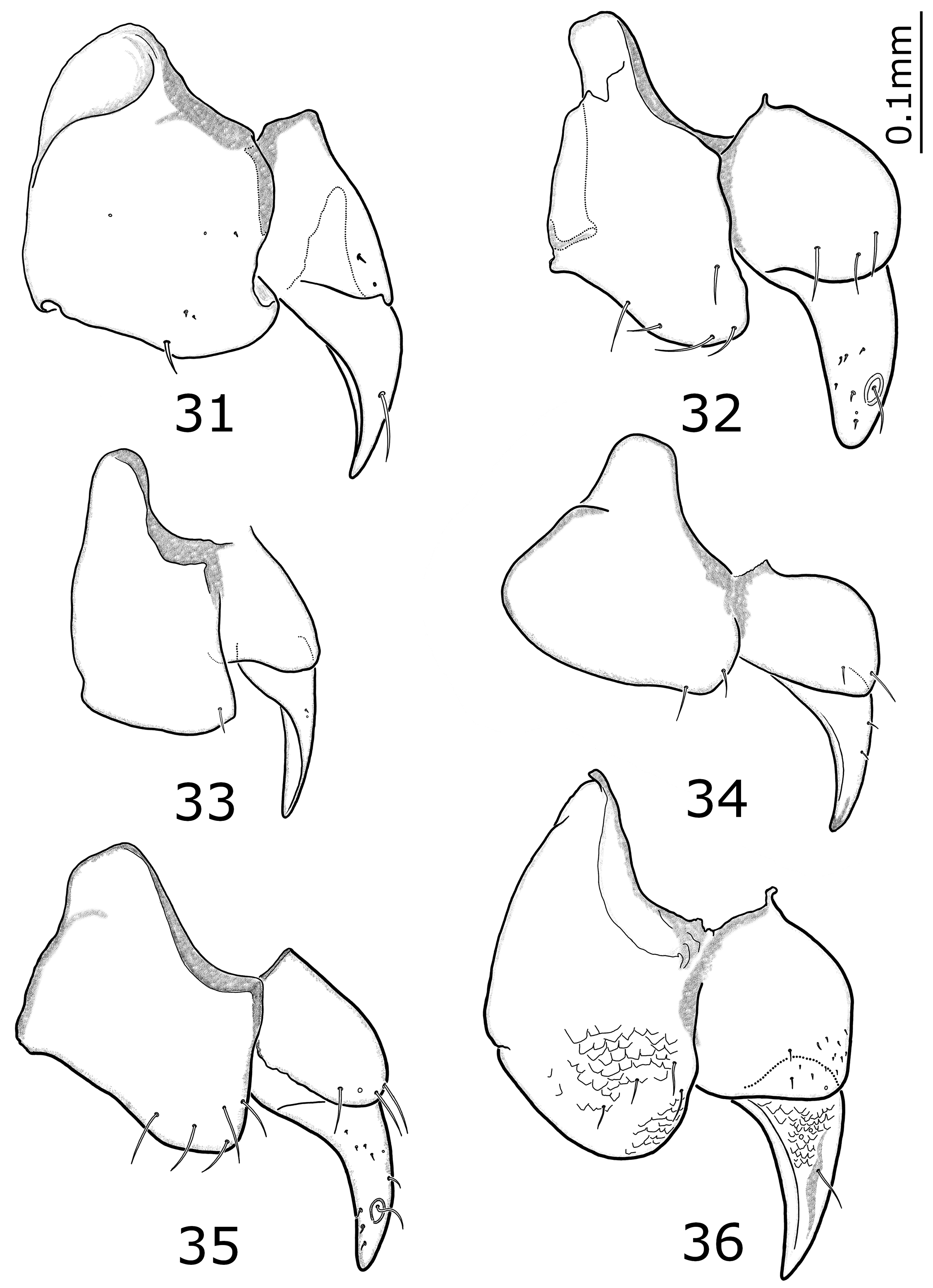
Female genitalia ( Figs. 31–36 View FIGURES 31 – 36 ): examination was possible only in six species: zupcense , kevser , albanicus sp. nov., jasminkoi sp. nov., rumijaensis sp. nov. and mlejneki sp. nov.. Gonocoxite 1 with one to three short setae close to the distal margin ( kevser , jasminkoi sp. nov., rumijaensis sp. nov.), or without them ( zupcense , albanicus sp. nov., mlejneki sp. nov.), gonocoxite 2 short, at apex rounded ( kevser ) or more or less pointed (other species), with two–ten very short setae ( jasminkoi , kevser , rumijaensis sp. nov.) or without them ( zupcense , albanicus sp. nov., mlejneki sp. nov.); sternal sensorial fovea small, with one long seta ( zupcense , kevser , rumijaensis sp. nov., mlejneki sp. nov.) or this seta is missing ( albanicus sp. nov., jasminkoi sp. nov.). We suppose that in older specimen examined (old dry preparation, specimen from long exposed traps) some especially short setae could be broken. This opinion should be confirmed by the examination of additional fresh specimen.
Geographical distribution. So far known from underground habitats (caves, pits) in Bosnia & Hercegovina, Montenegro and northern Albania (see Distribution map, Fig. 37 View FIGURE 37 ). Eight species are only known from a single limestone karst caves, four species ( pretneri , zupcense , mlejneki sp. nov. and rumijaensis sp. nov.) were found in more (two or three) underground localities. Adriaphaenops inhabits subterranean habitats situated from low altitudes ( pretneri from Vjetrenica cave on Popovo polje, ca 220 m a.s.l.) thorough middle regions up to highly situated caves ( kevser from Vilina pećina cave on Lebršnik Mts. , ca 1820 m a.s.l. and zupcense s.l. from Pećina u Zupcima cave, ca 2100 m a.s.l. and Jama na Vjetrenim Brdima, ca 2170 m a.s.l. on Durmitor Mts.). Low situated caves are warmer and mostly drier, with air temperature about 8–9 o C, mountain caves are significantly colder and wetter, with air temperature between 2–4 o C.
Bionomy. Virtually nothing is known about the bionomy of Adriaphaenops as well as other Dinaric genera of aphaenopsoid trechine beetles. Most of the taxa known so far, with the exception of Acheroniotes mlejneki Lohaj & Lakota, 2010 and Aphaenopsis apfelbecki (Ganglbauer, 1891) , were collected only individually or are known only from very few individuals. Representatives of these taxa were mostly found free walking or running on wet walls, speleothems and stones in the caves and pits, exceptionally collected also by pitfall traps baited with meat and/or cheese. Bionomy of Acheroniotes Lohaj & Lakota, 2010 is identical to other endogenous species of trechine genera like Duvalius Delarouzée , Duvaliopsis Jeannel or Trechus Clairville. No specimens were found outside of the clay, on the walls or stones. Special behaviour was observed in Scotoplanetes Absolon. Most of the known specimens were found in cave hygropetric environment ( Sket, 2004), in Vjetrenica cave (B&H) and Vodna jama pit ( Montenegro) together with leptodirine beetle Hadesia G. Müller.
Common external morphology. Strongly flattened body with very long sensorial setae on elytra and/or head and pronotum can indicate that these taxa are inhabitants of narrow spaces like fissures, slots, rock debris and MSS (Mesovoid Shallow Substratum or originally Milieu Souterrain Superficiel sensu Juberthie et al., 1980a, b, 1981). We suppose that conditions (temperature, humidity) and availability of food here is more stable than in large cave spaces, which they visited only occasionally. Morphology of mandibles, which are very long and slender, apically pointed and usually with robust teeth at the base of the right mandible, indicates that they are very active predators ( Casale et al., 2012; Lohaj & Lakota, 2010; Lakota et al., 2010; Mlejnek & Zajíček, 2006; Mlejnek, Jalžić, pers. comm.). Interestingly, ecomorphological convergent evolution between Dinaric Adriaphaenops and Pyrenean Hydraphaenops species (eg. H. vasconicus , H. navaricus , H. elhersi ...) provides evidences of adaptive processes as important drivers of phenotypic conservatism. Further research could develop these interesting topics.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Trechini |
Adriaphaenops Noesske, 1928
| Lohaj, Roman, Lakota, Ján, Quéinnec, Eric, Pavićević, Dragan & Čeplík, Dávid 2016 |
Adriaphaenops
| Moravec 2003: 289 |
| Casale 1994: 421 |
| Drovenik 1994: 43 |
| Monguzzi 1993: 238 |
Aphaenops
| Pavicevic 2001: 33 |
| Scheibel 1935: 34 |
Adriaphaenops
| Noesske 1928: 5 |
Aphaenopsis
| Pavicevic 1990: 365 |
| Vigna 1988: 166 |
| Casale 1982: 159 |
| Pretner 1959: 79 |
| Jeannel 1928: 793 |