
Vieja hartwegi ( Taylor & Miller 1980 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4375.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:31F06D99-E7DF-4A6E-A24A-781E98385ED6 |
|
DOI |
https://doi.org/10.5281/zenodo.5969912 |
|
persistent identifier |
https://treatment.plazi.org/id/03C0878B-FFE7-FFA1-E09D-4388FC2DC096 |
|
treatment provided by |
Plazi |
|
scientific name |
Vieja hartwegi ( Taylor & Miller 1980 ) |
| status |
|
Redescription of Vieja hartwegi ( Taylor & Miller 1980) View in CoL
Figs. 3–8 View FIGURE 3 View FIGURE 4 View FIGURE5 View FIGURE 6 View FIGURE 7 View FIGURE 8 , Tables 2, 3
Cichlasoma hartwegi Taylor & Miller 1980: 8 View in CoL [original description].
Paratheraps hartwegi ( Taylor & Miller 1980) View in CoL — Werner & Stawikowski 1987: 20 [ new combination].
Paraneetroplus hartwegi ( Taylor & Miller 1980) View in CoL — McMahan et al. 2010: 1298 [ new combination].
Material examined. All from Chiapas, Mexico, except FMNH 131457; numbers in parentheses indicate number of specimens examined. Vieja hartwegi (typical morph): UMMZ 207701 (holotype), Río Grande de Chiapa (= Grijalva River) about 1 km above bridge between Tuxtla Gutiérrez and Chiapa de Corzo; UMMZ 186400 (2 paratypes) Frío River, above mouth of Chiapa River, Acala; FMNH 93578 (1 paratype) Salado River, 1 km from Chiapilla; MZUNICACH 1053 (1), 1065 (2), 1125 (1), 1156 (1) Totopac River, Tecpatán; MZUNICACH 1560 (1), 1639 (1), 7158 (2) La Venta River, Ocozocoautla; MZUNICACH 1789 (2) Malpaso Reservoir, Ocozocoautla; MZUNICACH 3045 (3) Río Grijalva, Chicoasén; MZUNICACH 3145 (5) Grijalva River, Copainalá; MZUNICACH 5947 (1), 6245 (1) Santo Domingo River, Chiapa de Corzo; MZUNICACH 7053 (1) Chiquito River, Chiapa de Corzo; MZUNICACH 7153 (3) Yayahuita River, Chicomuselo; MZUNICACH 6761 (2 c&s), 7154 (8) La Angostura Reservoir, La Concordia; MZUNICACH 7208 (1) Copanó River, Ostuacán; ECOSC 7542 (2), 7544 (1) Blanco River, Venustiano Carranza; ECOSC 7543 (3), 7548 (3) Lagos de Colón, La Trinitaria; FMNH 131457 (2) Lagartero River, Nentón, Huehuetenango, Guatemala. Vieja hartwegi (rheophilic morph): MZUNICACH 7157 (1 + 1 c&s), 7160 (1), 7166 (1) Yayahuita River, Chicomuselo; MZUNICACH 7159 (1 + 1 c&s) Tachinua River, Chicomuselo; MZUNICACH 7161 (4) Lagos de Colón, La Trinitaria.
Diagnosis. The typical morph of V. hartwegi usually possesses a dorsolateral or second stripe thinner and horizontal (parallel) to the midlateral stripe ( Fig. 3 View FIGURE 3 B-C); in contrast, V. bifasciata ( Fig. 9A View FIGURE 9 ) has a broader second stripe that is slightly angled dorsally. The two stripes are separated by a narrow space (two thirds or half of a scale) or are often fused in V. hartwegi ( Fig. 5 View FIGURE5 D-J), whereas in V. bifasciata the space is broader (one and half or two scales). Vieja bifasciata usually has a large dark opercular blotch, extending to the border of the eye ( Fig. 9A View FIGURE 9 ), whereas V. hartwegi has a simple crescent shape ( Fig. 3 View FIGURE 3 B-C). Additionally, V. bifasciata has a much deeper body with a more rounded shape than V. hartwegi .
Vieja breidohri can be distinguished from V. hartwegi by the presence of molariform or subconical pharyngeal teeth, versus conical. Additionally, lateral blotches or a second stripe above the midlateral stripe are absent in V. breidohri ( Fig. 9B View FIGURE 9 ). The midlateral stripe in V. breidohri is markedly disrupted, formed by a series of separated or connected blotches that fade anteriorly, not reaching the opercle ( Fig. 9B View FIGURE 9 ). Juvenile specimens of V. hartwegi often possess a disrupted midlateral stripe below or at the middle of the body; in this case the two species are more difficult to distinguish. Vieja hartwegi possesses red spots or blotches on head ( Fig. 3 View FIGURE 3 A-C), whereas V. breidohri has small dark dots on sides of the head ( Fig. 9B View FIGURE 9 ). Vieja hartwegi possesses 29–31 total vertebrae (15 precaudal + 14–16 caudal), and V. breidohri has 28 (14 precaudal + 14 caudal).
All other species of Vieja with a midlateral stripe along the side of the body ( V. fenestrata , V. guttulata , and V. zonata ) can be differentiated from V. hartwegi by the absence of the second or dorsolateral stripe.
The rheophilic V. hartwegi morph differs from the typical morph and all other species of Vieja by a more elongated body shape ( Figs. 4 View FIGURE 4 & 6 View FIGURE 6 ), which contrasts with a deeper body in the typical morph of V. hartwegi and other species of Vieja . The body shape of specimens is similar to that of other rheophilic herichthyine cichlid species, such as Paraneetroplus and Rheoheros . The rheophilic morph also differs from all members of Vieja by the presence of a simple great spot ( Fig. 4 View FIGURE 4 A-C) or an ocellus ( Fig. 4D View FIGURE 4 ) at the center of the base of the caudal peduncle and on the origin of the caudal fin; other species of Vieja have a simple caudal spot ( V. maculicauda ) or the caudal spot is fused with the midlateral stripe (the rest of the species; e.g. V. melanura ). Also, in the rheophilic morph, the midlateral stripe is absent; only a midlateral blotch on the fifth bar is noticeable. This coloration pattern is similar to the syntopic Chiapaheros grammodes but can be differentiated by the great spot or ocellus on the caudal peduncle of the rheophilic V. hartwegi morph ( Fig. 6 View FIGURE 6 ) versus the simple oblong blotch in C. grammodes . This difference is always noticeable between juvenile and adult specimens. The midlateral blotch in C. grammodes is on the sixth bar versus the fifth bar in the rheophilic V. hartwegi morph. Chiapaheros grammodes has a series of thin brownish lines across interorbital region, snout and cheeks (the main character to diagnose this monotypic genus); in contrast, the rheophilic morph possesses two dark interorbital bars. Additionally, C. grammodes differs notably from the genus Vieja by the large head and enlarged snout, a large mouth with prognathous lower jaw. In preserved specimens, the rheophilic morph possesses a horizontal stripe above the middle of the body ( Fig. 6 View FIGURE 6 ), and in live specimens the band is noticeable under stress conditions ( Fig. 4 View FIGURE 4 B-D). This stripe is similar to the dorsolateral or second stripe of V. hartwegi and V. bifasciata , formed by the fusion of lateral blotches and always separated from the caudal blotch ( Fig. 6 View FIGURE 6 ). Other members of Vieja possess a strong midlateral stripe at level of mid-body ( V. breidohri ), below mid-body ( V. bifasciata , V. fenestrata , V. guttulata , V. zonata ), or variable, as in the typical V. hartwegi morph ( Figs. 3 View FIGURE 3 & 5 View FIGURE5 ).
Description. Morphometric and meristic data summarized in Tables 1–2. Maximum size 340 mm SL with weight of 1028 g in Grijalva River main channel, municipality of Chicoasén, Chiapas (observation from fishery La Cuevita). Meristics for the typical V. hartwegi morph ( Figs. 3 View FIGURE 3 & 5 View FIGURE5 ): Fins: D XVI–XVIII (mode 17), 11–14 (mode 12); A VI–VII (mode 6), 8–10 (mode 9); pectoral rays 15–17 (mode 16). Scales: Lateral series 30–34 (mode 32); upper lateral line 16–23 (mode 20); lower lateral line 8–15 (mode 12); in one specimen, pored scales of upper lateral line disrupted by non-pored scales (MZUNICACH 7158, specimen 2); subsidiary pored scales on caudal fin, usually 0–3 (mode 2) continuous with lower lateral line or interrupted by one non-pored scale; isolated pored scales (1–3) on the caudal-fin base, usually 1–2 scales above or below the subsidiary pored scales; predorsal squamation pattern irregular; scale row overlap 2–4 (mode 2); pectoral-pelvic scale rows 6–8 (mode 7); scale rows from lateral line to base of first dorsal-fin ray (not including the scaly sheet along the base), 3–4 1/2 (mode 3 1/2); scale rows from lateral line to base of first dorsal-fin spine 5–7 (mode 6); scale rows from origin of anal fin to lower lateral line 8–10 (mode 9); circumpeduncular scales 16–22 (mode 20); scale rows on cheek 5–7 (mode 5), one specimen with irregular pattern, not forming rows (ECOSC 7543, specimen 1). Cheek and opercle fully scaled, scales cycloid; dorsal portion of head and predorsal scales cycloid; base of dorsal and anal fins and scaly sheath along fin bases cycloid; lateral body scales ctenoid; lateral chest scales ctenoid, smaller than lateral body scales; ventral chest scales cycloid.
Body shape moderately deep or slightly slender ( Figs. 3 View FIGURE 3 & 5 View FIGURE5 ), in very large adults (> 200 mm SL) body deeper ( Fig. 3C View FIGURE 3 ); laterally compresssed; predorsal contour steep, dorsal head profile in front of orbit usually convex in juveniles, straight or slightly rounded in adults; the individuals with elongated snout, concavity present over the eyes; prepelvic contour gently convex; caudal peduncle short, deeper than long. Mouth small, usually low in position in juveniles and preadults, terminal and slightly oblique in large adults; individuals of upper reaches with more slender bodies, mouth usually terminal ( Figs. 5 View FIGURE5 F-G). Upper jaw in juveniles and preadults usually projected beyond lower jaw ( Fig. 5 View FIGURE5 A-D), large adults isognathous ( Fig. 5 View FIGURE5 G-J). Snout moderately pronounced or sometimes flattish. Lips of both jaws moderately thickened. Frenum well developed. Maxillary cleft position, variable but usually at level of dorsal margin of pectoral fin base.
Gillrakers on first arch: lower limb 7–11 (mode 9), one specimen found in sulphidic environment possessed only two gillrakers (MZUNICACH 1065, specimen 1); upper limb 1–3 (mode 2); total 9–13 (mode 11), one specimen with only 5 (see line above). Gillrakers moderately elongate, shorter and thickened in adults and more elongated in juveniles; tips pointed or rounded, commonly anterior gillrakers on lower limb shorter and rounded and posterior ones larger and pointed. One specimen, with posterior gillrakers bifurcated on lower limb (MZUNICACH 3145, specimen 1). Outer teeth in both jaws recurved, conical; anterior teeth enlarged with small posterior cusp; 3–4 irregular series of inner smaller teeth.
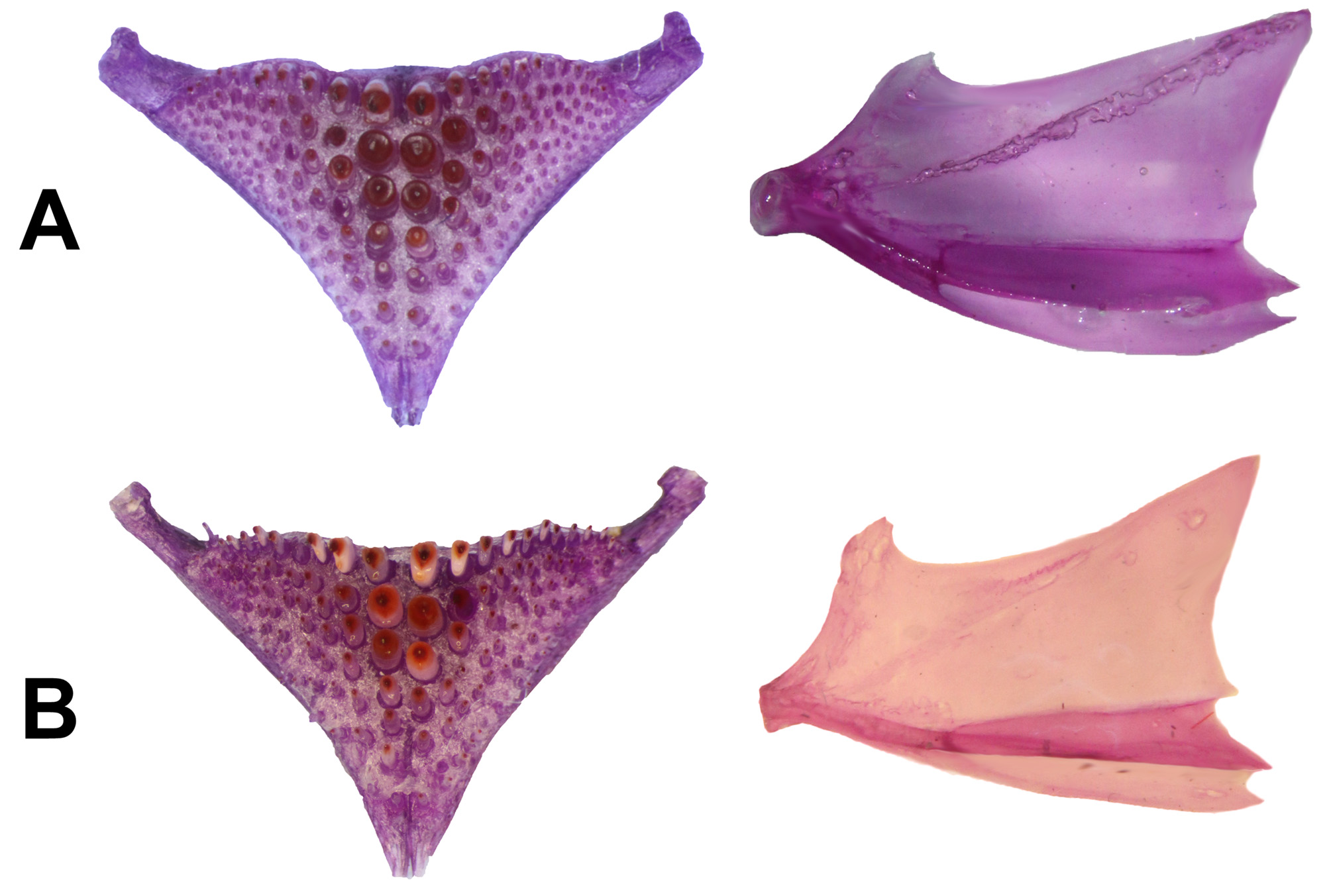
Lower pharyngeal tooth-plate ( Fig. 7A View FIGURE 7 ), dissected from one specimen (MZUNICACH 6761, 89.3 mm SL), dorsoventrally compressed, with short anterior process and posterior processes slightly enlarged; wider than long; posterior margin concave at center and convex at margins; teeth conical, enlarged posteriorly and shorter anteriorly, with antrorse cusp; second cusp small present at the base; three medial rows on each side, mainly the first, notably bigger and more robust posteriorly than the rest; 26 total (12/ 14 in each side) teeth in posterior row; 7 and 6 teeth in each near median row. Urohyal bone with posterior edge concave, dorsally slight convex with a short, curved spine ( Fig. 7A View FIGURE 7 ). Axial skeleton, based on 2 c&s specimens: vertebral counts, 15 precaudal and 14–15 caudal, total 29–30; ribs, 9 epipleurals and 13 pleurals; 2 supraneurals, between the supraoccipital crest and first dorsal pterygiophore; pterygiophores, 27 dorsal and 12–13 anal; the first anal pterygiophore contains 2 or 3 spines ( Fig. 8A View FIGURE 8 ).
Rheophilic morph smaller, largest known 97.1 mm SL, with following meristic data: Fins: D XVIII, 11–13 (mode 11), A V–VII (mode 6), 8–10 (mode 9); pectoral rays 15–16 (mode 16). Scales: Lateral series 31–34 (mode 31); upper lateral line 20–21 (mode 21); lower lateral line 9–13 (mode 12), in two cases, pored scales disrupted by non-pored scales and usually 1–3 (mode 2) subsidiary pored scales on caudal fin; isolated pored scales on caudalfin base, absent; predorsal squamation pattern irregular; scale row overlap 1–3 (mode 2); pectoral-pelvic scale rows 7–8 (mode 7); scale rows from lateral line to base of first dorsal-fin ray 3–3 1/2 (mode 3 1/2); scale rows from lateral line to base of first dorsal-fin spine 6–7 (mode 6); scale rows from origin of anal fin to lower lateral line 8–9 (mode 9); circumpeduncular scales 18–21 (mode 20); scale rows on cheek 6–7 (mode 7). Squamation pattern similar to typical morph.
Body shape slender or moderately deep ( Figs. 4 View FIGURE 4 & 6 View FIGURE 6 ); predorsal contour slightly steep, dorsal head profile in front of orbit usually straight or slightly convex; prepelvic contour gently convex; caudal peduncle short, deeper than long. Mouth small, terminal and isognathous in all sizes. Snout moderately pronunced. Lips of both jaws moderately thickened. Frenum well developed. Maxillary cleft position, usually ventral or sometimes at level of dorsal margin of the pectoral-fin base.
Gillrakers on first arch: lower limb 8–10 (mode 9); upper limb 3–4 (mode 3); total 11–13 (mode 13). Gillrakers moderately elongated; tips pointed or rounded, commonly anterior gillrakers on lower limb shorter and rounded, and posterior larger and pointed. Outer teeth in both jaws recurved and longer than typical morph, with posterior little cusp, anteriorly increasing in size; 3–4 irregular series of inner smaller teeth.
Lower pharyngeal tooth-plate ( Fig. 7B View FIGURE 7 ) dissected from two specimens (MZUNICACH 7157, 61.2 mm SL; 7159, 53.3 mm SL), dorsoventrally compressed, with short anterior process and posterior processes slightly enlarged; wider than large; posterior margin concave at center and very convex at margins; teeth conical, enlarged posteriorly and shorter anteriorly, with antrorse cusp; second cusp small but strong, present at the base; four or three medial rows, mainly the first, notably larger and more robust posteriorly than the rest; 22–24 (11/11 and 12/ 12 in each side) total teeth in posterior row; 8/7 and 6/7 teeth in near median rows. Urohyal bone with posterior edge concave, dorsally convex with a strong curved spine ( Fig. 7B View FIGURE 7 ). Axial skeleton, based on two c&s specimens and one radiograph: vertebral counts, 15 precaudal and 15(1)–16(2) caudal, total 30(1)–31(2); ribs, 12 epipleurals and 13 pleurals; 2 supraneurals, between the supraoccipital crest and first dorsal pterygiophore; pterygiophores, 30 dorsals and 15 anals; the first anal pterygiophore contains 2 spines ( Fig. 8B View FIGURE 8 ).
Coloration in preserved specimens. Vieja hartwegi has grayish background color, turning to brownish depending on time since preservation. Head usually somewhat darker than body. Ventral portion uniformly lighter. Typical morph with distinctive midlateral stripe extending from caudal-fin base to opercle, running at level ( Fig. 5A View FIGURE5 , C-D) or usually below mid-body ( Fig. 5B View FIGURE5 , E-J); a well-developed dorsolateral or second stripe slightly separated ( Fig. 5E, G, J View FIGURE5 ), partially fusioned anteriorly ( Fig. 5H View FIGURE5 ) or sometimes fully fused ( Fig. 5D, I View FIGURE5 ) with midlateral stripe in adults. In adults, second stripe formed by the fusion of blotches. In juveniles and subadults, midlateral stripe usually disrupted; commonly three to five irregular blotches above midlateral stripe ( Fig. 5 View FIGURE5 B-C, F) or rarely absent in some specimens; in small juveniles, cross-bars arranged along midlateral stripe. Adults in breeding coloration presents five cross-bars along midlateral stripe, lower jaw and belly darker (as the holotype, Fig. 5A View FIGURE5 ). Interorbital bars commonly not evident in fixed specimens. Black crescent-shaped operculum marking present in adults, in juveniles and preadults usually weak or absent. In some specimens, brown spots present on cheeks. Soft portions of dorsal and anal fins and caudal fin, dark brown spotted in both morphs.
Rheophilic morph with distinctive dorsolateral stripe present above of the middle of the body, extending from margin of the opercle to the level of dorsal fin posterior end ( Fig. 6A View FIGURE 6 ); this stripe is similar to dorsolateral or second stripe in typical morph, formed by the fusion of lateral blotches. Great rounded spot at center of caudal peduncle and base of caudal fin, often disrupted with dorsolateral stripe or sometimes confluent, but always remarkable ( Fig. 6 View FIGURE 6 ). Eight diffuse bars along the body, usually six bars notable, especially in juveniles ( Fig. 6B View FIGURE 6 ). Usually, series of 4–6 dark dots below dorsolateral stripe.
Coloration in life. Vieja hartwegi exhibits a highly variable coloration throughout its distributional range. Populations in upper Grijalva show background color that varies from silver-gray to olive-gray; in middle and lower Grijalva it varies from olive-gray to yellowish (see Conkel 1993: 58). Head dorsally olive-gray to yellowish; in juveniles and sub-adults red spots present on preoperculum, operculum, cheek, and lachrymal ( Fig. 3A View FIGURE 3 ); in adults red blotches cover entire head ( Fig. 3 View FIGURE 3 B-C). Side of head bluish and iridescent base color, more vibrant in sub-adults and adults. Two dark interorbital bars present. In large adults red dots at the center of the scales cover all of the body continuing onto dorsal, anal, and caudal fins ( Fig. 3C View FIGURE 3 ). Belly usually red in adults. In juveniles and subadults, dots along the body brownish red. Midlateral and dorsolateral (or blotches) stripes black; sometimes the two bars fully fused. Pectoral fins transparent or slightly yellowish; pelvic fins grayish. Dorsal and anal fins gray translucent in juveniles and sub-adults and yellowish in adults. In very large adults, spiny portion of the dorsal fin, golden. Border of dorsal fin red and anal fin dark. Caudal fin gray and translucent, with red border in large adults ( Fig. 3C View FIGURE 3 ). Breeding coloration is shown in Werner and Stawikowski (1987: 23).
Rheophilic morph has olive gray base color. Head olive gray dorsally with red or brownish orange spots or blotches. Two dark interorbital bars evident. Side of head bluish and iridescent, covered by variable brownish orange or reddish spots or blotches (usually forming a reticular pattern) all over opercle, preopercle, cheek, and lachrymal ( Fig. 4 View FIGURE 4 ). In some specimens, bluish iridescent horizontal stripe prominent below eye ( Fig. 4C View FIGURE 4 ). In normal coloration, two dark marks notable: a great rounded spot at center of the caudal peduncle and base of caudal fin ( Fig. 4 View FIGURE 4 A-C; some specimens present an ocellus, with iridescent light blue around the spot, probably a sexual dimorfism in males Fig. 4D View FIGURE 4 ) and irregular blotch on the middle of the body. Under stress, coloration consists of dark vertical bars along the body and series of seven blotches along the bars, forming a general dorsolateral stripe above the middle of the body with no fusion of caudal spot or ocellus ( Fig. 4 View FIGURE 4 B-D). Distal portions of lateral scales reddish or brownish orange, whereas proximal portions iridescent light blue, forming regular series of longitudinal stripes along each scale row. Pectoral fins transparent or slightly yellowish; pelvic fins pale or dark grayish. Dorsal and anal fins brownish or yellowish and translucent with bluish iridescent dots. Border of dorsal fin red with bluish line below the red and across the spiny portion of the fin. Border of anal fin dark; caudal fin gray and translucent with regular series of brownish and sometimes bluish iridescent dots.
Distribution and habitat. Found on the Atlantic slope of Chiapas, Mexico and Guatemala, endemic to the Grijalva River basin. Miller et al. (2005) considered its distribution only in the middle and upper portions of the Grijalva. Gómez-González et al. (2015) documented its presence in the lower Grijalva in Ángel Albino Corzo or Peñitas reservoir and tributaries. It also inhabits the upper reaches of the lower Grijalva outside of reservoirs ( Fig. 10 View FIGURE 10 ). Elevations typically range from 100 to 700 m; however, an exceptional record exists from 1600 m in the isolated Laguna Verde, municipality of Coapilla, Chiapas (MZUNICACH 1239, 1242). The rheophilic morph is restricted to tributaries of the far upper Grijalva River basin, only known in Lagartero River, near the Guatemalan border, and the Tachinua and Yayahuita rivers in Chiapas, Mexico ( Fig. 10 View FIGURE 10 ).
The typical V. hartwegi morph inhabits a great variety of habitats. Large adults prefer deeper zones of the main river channel (> 2 m depth) with variable but mainly moderate current and rocky or sandy substrates, usually without aquatic vegetation. Juveniles and subadults prefer shallow waters with rocky bottoms, sand, silt and mud in rivers and streams with swift to slow currents, usually without aquatic vegetation but covered by riverine vegetation. Also inhabits lakes and reservoirs. The species is also tolerant to sulphidic environments; it has been recorded in El Azufre and Baños del Carmen, municipalities of Tecpatán and Venustiano Carranza, respectively, both in Chiapas, Mexico.
The rheophilic morph primarily inhabits rocky bottoms covered with boulders and rocks of various sizes and wooden debris, aquatic vegetation absent, in depths of 30–100 cm; moderate to fast currents, with clear or slightly murky waters. This morph of V. hartwegi is sympatric with the typical morph; however, the two forms are likely segregated because of habitat preferences and food habits. There are too few specimens to robustly analyze the food preferences, but slight differences in pharyngeal jaw dentition may be related to diet. Additionally, V. breidohri and C. grammodes are found in sympatry with this morph.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Vieja hartwegi ( Taylor & Miller 1980 )
| Gómez-González, Adán E., Álvarez, Fernando, Matamoros, Wilfredo A., Velázquez-Velázquez, Ernesto, Schmitter-Soto, Juan J., González-Díaz, Alfonso A. & Mcmahan, Caleb D. 2018 |
Cichlasoma hartwegi
| Taylor & Miller 1980: 8 |