
Bilyjomyia fontana, Niitsuma, Hiromi & Watson, Charles N., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.189110 |
|
DOI |
https://doi.org/10.5281/zenodo.5631218 |
|
persistent identifier |
https://treatment.plazi.org/id/03C05703-FFF6-4933-FF3E-FD67C5EEFBFA |
|
treatment provided by |
Plazi |
|
scientific name |
Bilyjomyia fontana |
| status |
sp. nov. |
Bilyjomyia fontana View in CoL new species
( Figs. 1 View FIGURES 1 – 5 –21)
Type material. Holotype male with its pupal and larval exuviae (SUM-IC-T0254). Japan: Fukushima Prefecture, Iwaki City, spring in Yaguki, 18.iii.2007 (adult emerged 25.iii.2007), leg. H. Niitsuma. Paratypes: 8 larvae (SUM-IC-T0255–0262), same data as holotype except 2.i.2007; 3 males with their pupal and larval exuviae (SUM-IC-T0263–0265), 4 females with their pupal exuviae and 3 of their larval exuviae (SUM-IC- T0266–0269), 2 pupae with their larval exuviae (SUM-IC-T0270–0271), 8 larvae (SUM-IC-T0272–0279), same data as holotype (these adults emerged 26.iii–10.iv.2007); 1 larva (SUM-IC-T0280), same data as holotype except 19.viii.2007; 1 female with its pupal exuviae (SUM-IC-T0281), 2 pupae (SUM-IC- T0282–0283), 1 larva (SUM-IC-T0284), same data as holotype except 4.v.2008 (adult emerged 8.v.2008).
Etymology. From the Latin fontanus (spring), referring to the habitat of the larvae.
Description. Male (n = 4). Body length 4.8–5.0 mm.
Coloration. Head yellowish-brown. Thorax generally yellowish-brown, with dark brown antepronotum, median anepisternum II, preepisternum and postnotum; scutal vittae slightly darker than rest of scutum. Wing with single dark band as in Fig. 1 View FIGURES 1 – 5 , formed by black macrotrichiae and light infuscation on membrane. Legs generally yellowish; femur reddish-brown on basal third; ta5 entirely dark brown. Abdominal tergites I–IV yellowish; T.V–VIII dark brown. Hypopygium dark brown.
Head. Temporals 42–46, multiserial. AR 1.9–2.0. Clypeus trapezoidal with 29–36 setae. Lengths of palpomeres 1–5 (μm) 50–60, 58; 75–85, 78; 180–213, 198; 220–300, 254; 275–363, 331. Pm5 1.2–1.4 times as long as Pm4.
Thorax. Antepronotals 18–23. Acrostichals about 100–120; dorsocentrals about 70–80; humerals about 30–40; supra-alars 47–50; postalars 1; pteropleurals 8–12; mesosternals 3–6. Scutellars 64–76, multiserial. Postnotum with 6–11 dorsomedial setae, biserial.
Wing ( Fig. 1 View FIGURES 1 – 5 ). Length 3.2–3.3 mm. Membrane with dense macrotrichae; VR 0.90. Squama with 39–45 setae.
Legs. Spur of foretibia 73–80 μm long with 16–18 lateral teeth; spurs of midtibia 85–95 and 55–63 μm long each with 16–18 lateral teeth; spurs of hind tibia 85–100 and 50–55 μm long each with 15 lateral teeth; comb of hind tibia consisting of 8–10 spines. Foreleg with short tarsal beard. Claw pointed apically with 2 or 3 basoventral spines. Lengths and proportions of leg segments as in Table 1 View TABLE 1 .
Hypopygium ( Fig. 2 View FIGURES 1 – 5 ). Tergite IX with 1–8 posterior setae. Gonocoxite 225–245 μm long, 2.3–2.5 times as long as broad in middle. Gonostylus 138–145 μm long with 1–3 apical megasetae ( Fig. 3 View FIGURES 1 – 5 ); HR 1.6–1.7. Phallapodeme straight or slightly curved.
Female (n = 5). Body length 3.8–4.2 mm.
Coloration. Similar to male, except T.III–VII entirely dark brown. Wing pattern ( Fig. 4 View FIGURES 1 – 5 ) more extensive, with anal cell infuscate.
Head. Temporals 42–54. Antenna with terminal flagellomere almost as long as preceding 3 segments together; AR 0.23–0.27. Clypeus with 35–39 setae. Lengths of palpomeres 1–5 (μm) 60–75, 68;75–95, 86; 220–240, 231; 305–355, 327; 350–430, 403. Pm5 1.1–1.3 times as long as Pm4.
(n = 3).
Thorax. Antepronotals 38–63; acrostichals about 130–160; dorsocentrals about 80–100; humerals about 70–90; supra-alars 61–74; postalars 1–2, usually 1; pteropleurals 8–18; mesosternals 9–15; scutellars 66–93; postnotals 6–15.
Wing ( Fig. 4 View FIGURES 1 – 5 ). Length 3.3–3.9 mm. VR 0.90–0.91. Squama with 77–83 setae.
Legs. Spur of foretibia 75–88 μm long with 16–18 lateral teeth; spurs of midtibia 93–105 and 55–65 μm long each with 16–18 lateral teeth; spurs of hind tibia 95–105 and 45–55 μm long with 14–18 and 10–17 lateral teeth, respectively; comb of hind tibia consisting of 8–10 spines. Midleg with 10–14 sensilla chaetica on apical 0.1 of ta1, and hind leg with 12–16 sensilla chaetica on apical 0.1 of ta1. Claw pointed apically with 3–4 basoventral spines. Lengths and proportions of leg segments as in Table 2 View TABLE 2 .
female (n = 5).
Genitalia ( Fig. 5 View FIGURES 1 – 5 ). Notum 145–175 μm long, 1.8–2.0 times as long as rami. Segment X with 0–2 setae on each side. Seminal capsule ovoid, 75–78 μm long, 53–55 μm wide, and slightly infuscated on apical 2/3; neck region pale. Cercus 63–90 μm long.
Pupa (n = 13). Body length 6.0– 7.2 mm.
Coloration. Exuviae generally brown; T.II–VII usually pale around bases of D-setae. Thoracic horn light brown.
Cephalothorax. Thoracic horn (Fig. 6) 400–500 μm long, 1.6–1.8 times as long as its broadest width, with 37–51 ostia on apical margin. Plastron plate 160–240 μm long, 0.68–0.81 times as long as broad, occupying distal 0.38–0.50 of horn; median aeropyle occasionally indistinct; lateral aeropyle located equidistant from median aeropyle and basolateral corner of plastron plate, and its opening elongate and diffuse. Thoracic seta Dc 1 30–38 μm long, 0.41–0.60 times as long as Sa; Dc2 2–4 μm long (Fig. 7).
Abdomen (Fig. 8). Scar of T. I 65 –115 μm long, 0.19–0.33 times as long as tergite along median line (Fig. 9). Shagreen mainly consisting of serial rows of 2–5 spinules (Fig. 10). Tergite IV with D1-seta 105–160 μm long, 0.22–0.28 times as long as segment; with D2- and D3-setae 330–380 μm long and 0.61–0.70 times as long as segment. LS1-seta of A.VII located 0.43–0.55 from anterior margin, of A.VIII located 0.61–0.72 from anterior margin. Anal lobe 909–1071 μm long, 2.6–2.8 times as long as broad, with anterior macroseta located 0.22–0.26 from anterior margin and posterior macroseta located 0.27–0.31; inner margin with 19–24 spinules on distal 0.42–0.53; outer margin nearly straight. Male genital sac 0.35–0.39 times as long as anal lobe.
Fourth instar larva (n = 27). Total length 7.1–11.6 mm.
Coloration. Body red when alive. Head generally reddish, and brown posterodorsally.
Head (Fig. 11). Length 909–1066 μm. IC 0.74–0.79. Labral sclerite (Fig. 12) amoebiform in shape; divided into several parts in some specimens; 0.14–0.20 times as wide as distance between two labral setae S2- S2 (W/D in Fig. 13). Cephalic seta S10 almost twice as long as S9. Antenna (Fig. 14) 1.2–1.4 times as long as mandible; lengths of segments 1–4 (μm, n =10) 248–288, 267; 15–20, 18; 4–5, 5; 4–5, 5. AR 7.7–8.7. First segment with ring organ placed 0.77–0.82 from base; blade 25–30 μm long, reaching middle of segment 4; accessory blade shorter than blade, 20–25 μm long. Second segment 2.9–3.3 times as long as wide; style 13–15 μm long, reaching segment 4; peg sensilla 8–10 μm long, reaching base of segment 3 (Fig. 15). Mandible (Fig. 16) 213–253 μm long. Basal segment of maxillary palp (Fig. 17) 68–75 μm long, 3.4–3.8 times as long as wide; ring organ located 0.23–0.28 from base. Dorsomental plate with 7–8 large teeth and usually 1 small tooth (Fig. 18). Ligula (Fig. 19) 128–145 μm long, 1.5–1.7 times as long as its toothed width; granulose area at base expanded medially, 0.35–0.42 times as long as its basal width; median tooth 1.6–1.9 times as long as wide; inner laterals pointed forward or turned slightly inwards; outer laterals 2.0–2.4 times as long as wide. Paraligula 63–70 μm long; outer tooth 9–14 times as long as inner tooth. Pecten hypopharyngis (Fig. 20) with 1 or 2 broadened mesal teeth, followed by row of 12–16 simple teeth becoming longer towards middle and 3–9 small points near bases of distal teeth.
Body. With lateral fringe of setae at most 0.3 times as long as segment width. Procercus 3.8–5.0 times as long as wide; both lateral setae located 0.28–0.37 and 0.79–0.85 from base, respectively. Posterior parapod with smaller claws depressed and expanded basally (Fig. 21).
Remarks. The adult of Bilyjomyia fontana differs from that of B. algens mostly in the coloration. The legs are brownish only on the basal part of femur and ta5. In Bilyjomyia algens the legs are brownish on the apex of femur, the base of tibia, and ta4, 5. The female abdomen is brown, and the male is dark brown on A.V–VIII; whereas, in the latter species the female abdomen is entirely pale, and the male is darkened on A.VI–VIII. The dark band of the female wing is expanded to encompass the entire anal cell. The anal cell of female Bilyjomyia algens is mostly pale (see Roback 1971, fig. 193). The male hypopygium is distinct, too, in the almost straight phallapodeme. In Bilyjomyia algens , the phallapodeme has a strong bend near the mesial end (see Roback 1971, fig. 188), that may be a unique feature among Macropelopiini .
The pupa is separable from that of Bilyjomyia algens by the thoracic horn with lateral aeropyles located nearly equidistant from the median aeropyle and the basolateral corner of the plastron plate, with elongate and diffuse openings. In Bilyjomyia algens the lateral aeropyles are closer to the basolateral corners of the plastron plate (see Roback 1984, figs. 2 and 3), and the openings are spherical and prominent. Furthermore, Bilyjomyia fontana has relatively short thoracic Dc1-setae, which are 0.41–0.60 times as long as the Sa-seta, and a poorly developed scar on T. I, which is 0.19–0.33 times as long as the tergite length along the median line. In Bilyjomyia algens the Dc1-setae are 0.71–0.88 times as long as the Sa-seta. The scar on T.I is well developed, and 0.32–0.45 times as long as the tergite length along the median line.
The larva of Bilyjomyia fontana differs from that of B. algens in the features of the labral sclerite which is relatively large; 0.14–0.20 times as wide as the distance between S2-S2; amoebiform in shape; with a tendency to divide into several parts. In the latter the sclerite is rounded and compact, 0.07–0.10 times as wide as the distance between S2-S2.
Ecology. Like most Macropelopiini , Bilyjomyia larvae are cold-stenothermal, confined to depositional substrates of springs, seeps, and headwater streams.
In Japan, Bilyjomyia fontana was collected with Macropelopia kibunensis (Tokunaga) and M. paranebulosa Fittkau , from a forest in Iwaki City, Fukushima Prefecture. The larvae inhabit a spring (13–15°C around noon in summer) with muddy bottom.
In North America, the immatures of Bilyjomyia algens have been collected from small, cold streams in the Pacific Northwest. In these streams, they are confined to the mud substrate of shallow pools ( Roback 1984, Watson 1998).
Biogeography. The distribution of Bilyjomyia in Asia and western North America follows the Beringian pattern, track 4 of the eight most common generalized biogeographic tracks discussed by Saether (2000). Among the Chironomidae , in addition to the taxa discussed by Saether (2000), this distribution pattern is also found in Brundiniella , Alotanypus , Compteromesa Saether , Antillocladius Saether , Compterosmittia Saether and Skutzia Reiss ( Niitsuma 2003, 2005; Niitsuma & Makarchenko 1997; Mendes et al. 2004; De Pinho et al. 2009). This pattern also occurs in other groups of insects. Peters (1988) reviewed the origins of the North American Ephemeroptera fauna, and noted that there were vicariant species pairs of mayflies in Asia and western North America. Sixteen of these genera are distributed primarily in western North America and the eastern Palearctic, a fact that they attributed to an extensive mid- to late-Pliocene trans-Bering exchange. Stewart & Stark (2002) found that the distributions of 36 genera of Plecoptera reflect a Bering Land Bridge dispersal or vicariance. Schmid (1970, cited in Saether 2000) found this pattern in Rhyacophila Pictet and other rheophilic Trichoptera .
The close similarity of Biljomyia algens and B. fontana suggests fairly recent dispersal or vicariance. Bilyjomyia must be present on the Asian mainland. Additional collecting is necessary to determine if there are additional species, and the geographic range of the genus. It would not be surprising to find additional species of Bilyjomyia , especially in poorly collected areas of Asia. Some taxa with an amphi-Pacific distribution range into south Asia, or even further. In the Nearctic, they may range southwards into Central and South America ( Saether 2000).
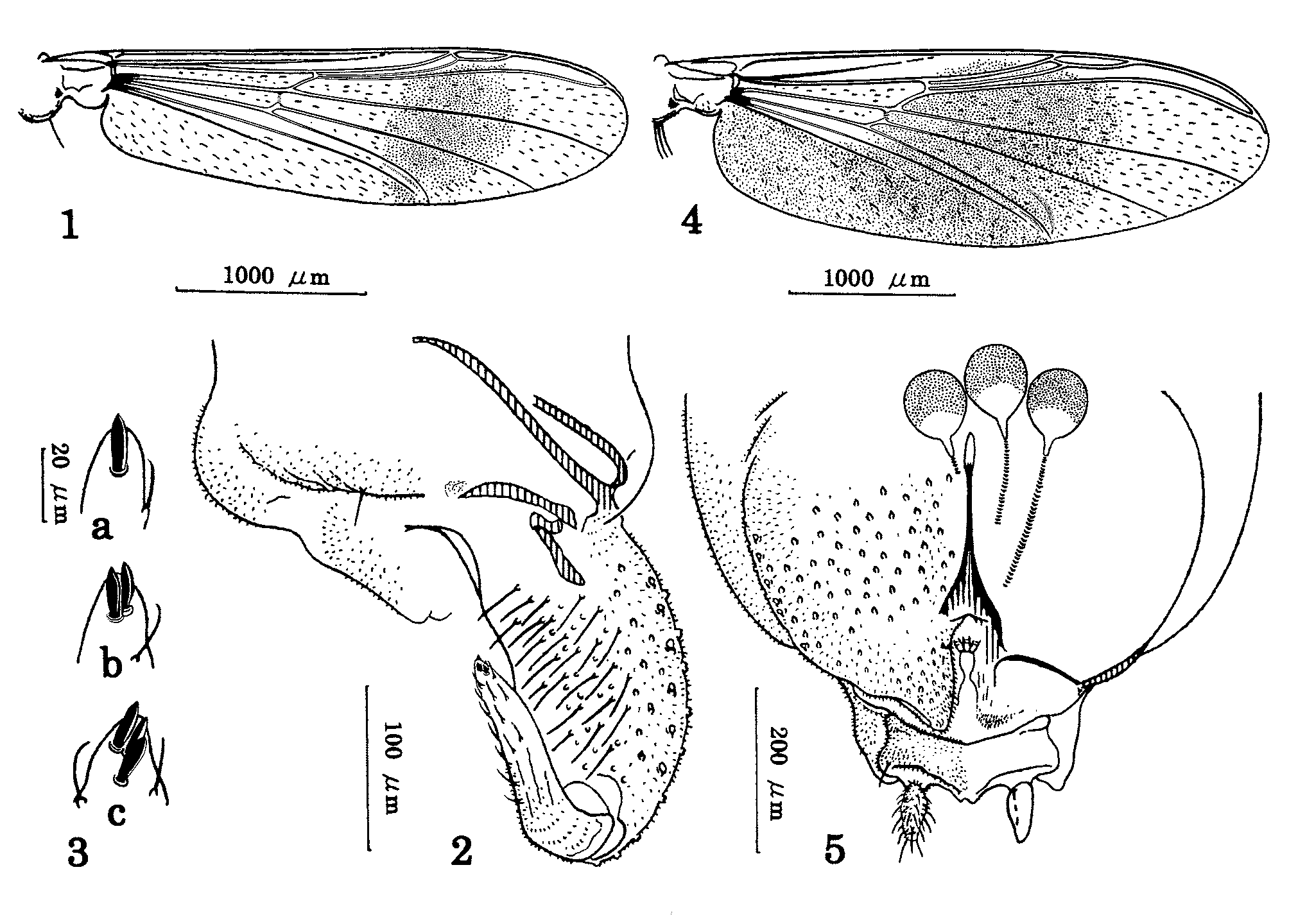
TABLE 1. Length (range and mean, in µm) and proportions of legs of Bilyjomyia fontana new genus, new species, male
| fe | ti | ta1 | ta2 | ta3 | ta4 | ta5 | LR | BR | |
|---|---|---|---|---|---|---|---|---|---|
| p1 | 1421–1447 1430 | 1624–1700 1675 | 1117–1269 1189 | 609–660 626 | 406–457 431 | 254–266 258 | 165–178 169 | 0.69–0.75 | 5.2–5.8 |
| p2 | 1421–1548 1497 | 1371–1523 1464 | 876–939 910 | 457–508 482 | 305–355 334 | 203, 203 | 127–140 136 | 0.61–0.64 | 4.8–5.8 |
| p3 | 1307–1421 1375 | 1624–1840 1764 | 1142–1345 1269 | 558–660 618 | 419–482 461 | 254–279 262 | 152, 152 | 0.70–0.73 | 5.9–6.0 |
TABLE 2. Length (range and mean, in µm) and proportions of legs of Bilyjomyia fontana new genus, new species,
| fe | ti | ta1 | ta2 | ta3 | ta4 | ta5 | LR |
|---|---|---|---|---|---|---|---|
| p1 1371–1675 1528 | 1726–2107 1939 | 1142–1371 1274 | 609–761 690 | 381–533 457 | 228–305 274 | 178, 178 | 0.63–0.69 |
| p2 1523–1878 1721 | 1459–1954 1761 | 964–1117 1061 | 533–584 563 | 355–457 411 | 216–279 251 | 140–168 153 | 0.56–0.66 |
| p3 1345–1675 1512 | 1802–2259 2056 | 1167–1421 1330 | 622–761 713 | 457–584 543 | 254–355 307 | 152–178 162 | 0.62–0.70 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Macropelopiini |
|
Genus |