
Trichomycterus maracaya, Bockmann & Sazima, 2004
|
publication ID |
https://doi.org/10.1590/S1679-62252004000200003 |
|
publication LSID |
lsid:zoobank.org:pub:84A1F738-EB50-4C16-957E-CEC257AF4F2F |
|
persistent identifier |
https://treatment.plazi.org/id/1F83BB4D-68B6-4DCC-B9D7-75BCBBFC34D0 |
|
taxon LSID |
lsid:zoobank.org:act:1F83BB4D-68B6-4DCC-B9D7-75BCBBFC34D0 |
|
treatment provided by |
Carolina |
|
scientific name |
Trichomycterus maracaya |
| status |
sp. nov. |
Trichomycterus maracaya View in CoL , new species Figs. 1-8 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig
Holotype. ZUEC 6137 View Materials , 48.8 mm SL; unsexed; Brazil, Minas Gerais, Poços de Caldas, rio Pardo basin, unnamed streamlet at the foothill of the Morro do Ferro (about 46º31’50”S, 21º46’45”W); C. F. B. Haddad & I. Sazima; 6 February 2003. GoogleMaps
Paratypes. 17 specimens: LIRP 4381 View Materials , 3 View Materials , 26.7-51.3 mm SL, 1 c&s, 43.9 mm SL; MCP 34575, 2 View Materials , 30.7-31.6 mm SL; MNRJ 25872 View Materials , 2 View Materials , 31.8-33.6 mm SL; MZUSP 83357 View Materials , 2 View Materials , 29.6-41.9 mm SL; ZUEC 6136 View Materials , 3 View Materials , 24.1-45.6 mm SL; ZUEC 6138 View Materials , 2 View Materials , 22.8-48.1 mm SL, 1 c&s, 32.5 mm SL; all collected with the holotype; ZUEC 4115 View Materials , 1 View Materials , 45.9 mm SL; same locality as holotype; A. J. Cardoso, C. F. B. Haddad & G. U. Andrade; 8 January 1981 GoogleMaps .
Diagnosis. Trichomycterus maracaya , along with T.brasiliensis , T. iheringi (Eigenmann) , T. mimonha Costa , T. potschi , and T. vermiculatus , are members of the Trichomycterus brasiliensis species-complex, based on the possession of the following apomorphic characters unique in Trichomycterus : four longitudinal rows of well-defined blotches formed by very dense concentration of dark chromatophores in the deeper layer of skin (vs. four rows faint, with limits of blotches poorly-defined); and pectoral fin with I+5-6 rays (vs. usually I+7-8). Members of this putative clade (except for T. iheringi ) may be further distinguished from other species of Trichomycterus in having the bases of the pelvic fins very close to each other, sometimes in contact (vs. the pelvic–fin bases very spaced). Additionally, T.maracaya differs from the remaining species of Trichomycterus (except T. brasiliensis and T. potschi ) in retaining the primordial epiphyseal cartilaginous bar separating the anterior and posterior cranial fontanels even in larger individuals (vs. having the osseous epiphyseal bar separating the anterior and posterior cranial fontanels appearing early in the ontogeny). Furthermore, T. maracaya is distinguished from other species of the T. brasiliensis species-complex, and from all congeners, in the following putative autapomorphies: 1) row of blotches on lateral surface not forming a lateral stripe in any stage during ontogeny (vs. lateral row of blotches, when present, forming a lateral stripe usually conspicuous early in ontogeny); 2) superficial layer of pigmentation of juveniles and larger, presumably adult, specimens formed by scattered chromatophores (vs. superficial layer of pigmentation, when present, of larger specimens formed by dense pattern of chromatophores often overlaying the larger blotches of the lower layer of pigmentation).
Description. Morphometric data for type series presented in Table 1. Dorsal profile of body sloped from tip of snout to anterior portion of trunk and approximately straight from that point to baseofcaudalfin ( Figs.1-3 View Fig View Fig View Fig ). Convexityoftrunkmorepronounced in larger individuals.Ventral body profile approximately straight to slightly convex from mouth to insertion of pelvic fin; slightly concave from end of base of pelvic fin to caudal fin. Crosssection of body in trunk region approximately oval, becoming gradually more compressed transversely posterior to base of dorsal fin. Caudal peduncle strongly compressed. Axillary gland with small pore. Urogenital and anal openings on vertical through base of first or second branched dorsal-fin rays.
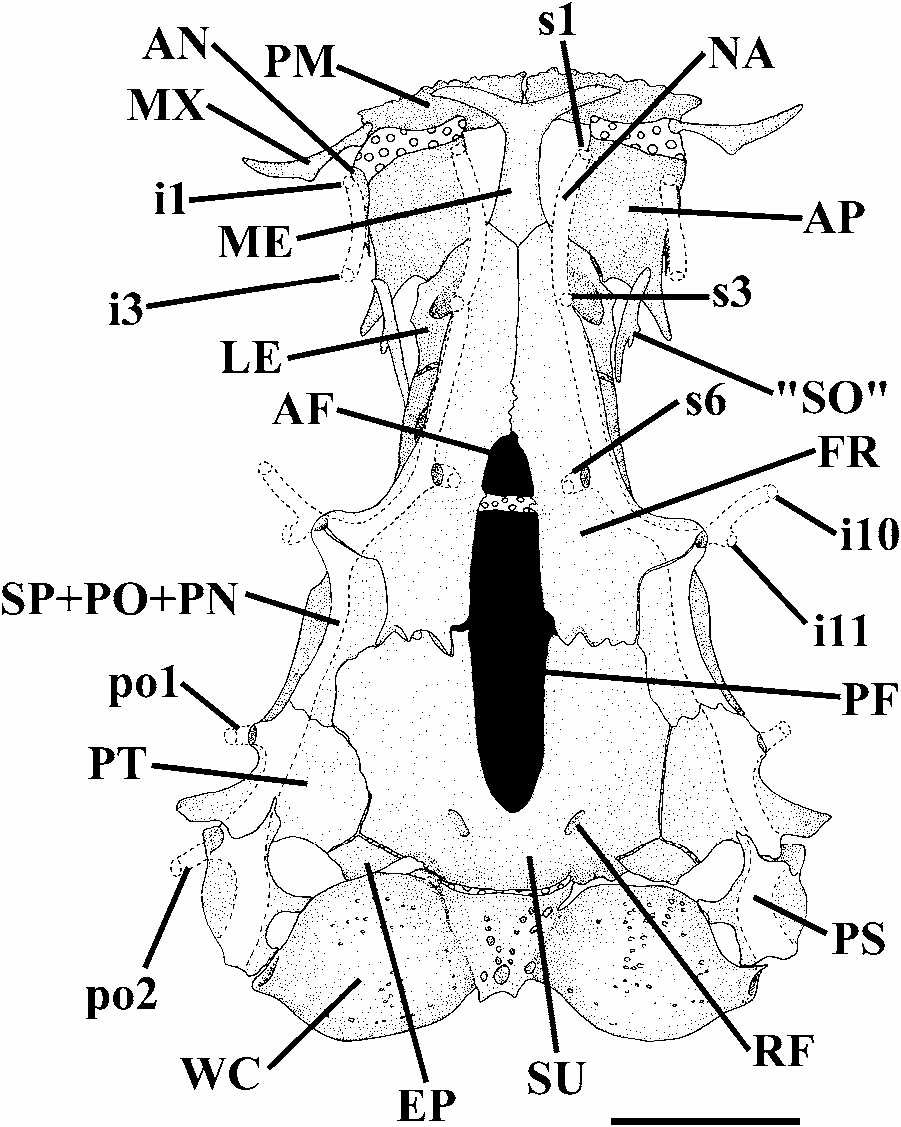
Head wide and depressed, slightly longer than broad ( Fig. 4 View Fig ). Eyes small, orbital rim not free except for discrete ventral invagination. Skin covering eye thin and translucent. Anterior nostril small, surrounded by fleshy tube-shaped flap of integument. Posterior nostril located approximately midway between anterior nostril and anterior orbital rim, and surrounded by raised fleshy flap, except posteriorly. Mouth inferior. Lower lip with conspicuous lateral fleshy lobes medial to origin of rictal barbels. Nasal barbel originating on posterolateral portion of integumentary flap around anterior nostril; tips of barbel reaching base of pectoral fin. Maxillary barbel extending to posterior border of pectoral-fin base. Rictal barbel shorter than maxillary barbel; tip reaching pectoral-fin base. Barbels progressively tapering distally. Opercular patch of odontodes small and elliptical. Interopercular patch of odontodes narrow, elongate and bearing conspicuous posterior projections but with anterior process very short. Interopercular with seven or eight odontodes in the smaller c&s specimen (32.5 mm SL) and 12-14 in the larger c&s specimen (43.9 mm SL). Opercle with 20 odontodes in the smaller specimen (32.5 mm SL) and 26 odontodes in the larger specimen (43.9 mm SL). Odontodes on both opercle and interopercle gradually increasing in size posteriorly. Tips of larger and posterior odontodes of opercle curved dorsomedially. Branchial membranes thick, united to isthmus only anteriorly and forming small free fold across isthmus. Branchiostegal rays hardly visible through thick skin.
Anterior cornua of mesethmoid with slight anterior concavity ( Fig. 5 View Fig ). Anterior fontanel restricted to small cuneiform opening situated immediately anterior to epiphyseal bar. Epiphyseal bar entirely cartilaginous, except for limited, medially-directed bony projection of frontals. Posterior cranial fontanel long and relatively broad between frontals and extending into parieto-supraoccipital. Antorbital short and anteriorly expanded. Tendon-bone supraorbital rod-like, approximately twice as long as antorbital. Anterior portion of sphenotic laterally directed in dorsal view. Sphenotic, prootic, and pterosphenotic entirely fused to each other. Vomer arrowshaped, with long posterior process. Parasphenoid with long and pointed posterior process that reaches posteromedian region of basioccipital. Anterior portion of Weberian complex fused to basioccipital. Weberian capsule with reduced lateral opening that is much smaller than lateral profile of capsule.
Premaxilla with 4 rows of conical teeth; anteriormost row with 12-14 teeth. Maxilla large, boomerang-shaped, and slightly shorter than premaxilla. Lower jaw with 3 rows of conical teeth, smaller c&s specimen (32.5 mm SL) with 11 teeth in most external row, and larger c&s specimen (43.9 mm SL) with 13-14 teeth in most external row.
Metapterygoid laminar and very large ( Fig. 6 View Fig ). Hyomandibula with conspicuous membranous outgrowth on its anterior margin. Articulation between metapterygoid and quadrate via anterior cartilage block only. Small, somewhat triangular preopercle sutures with ventral margins of both quadrate and hyomandibula.
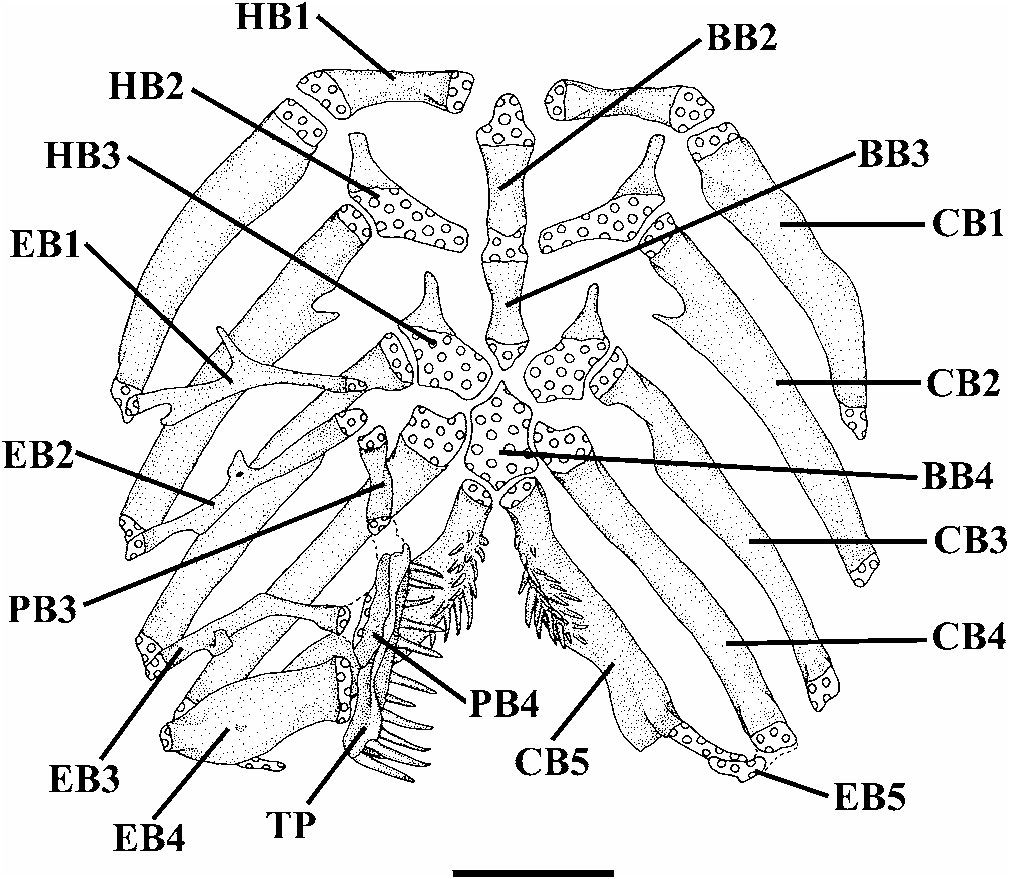
Hyoid arch with large ventral hypohyal, elongate anterior ceratohyal, and approximately short triangular posterior ceratohyal ( Fig. 7 View Fig ). Seven branchiostegal rays (1 specimen with anterior vestigial nodular ray not considered) articulated with hyoid arch: 2 with posteroventral margin of anterior ceratohyal, 1 with mesoventral margin of posterior ceratohyal, and 4 with cartilage between these bones. Branchiostegal rays 3-5 distally expanded and notched. Dorsal hypohyal and interhyal absent.
Urohyal with expanded anterior head, elongate lateral processes, and sharp and moderately short posterior process. Lateral processes of urohyal distally pointed. Basibranchial 1 absent. Basibranchials 2 and 3 with cartilaginous caps at their anterior and posterior tips, and connected to each other forming long rod ( Fig. 8 View Fig ). Anterior tip of basibranchial axis extending to level of hypobranchials 1, not reaching dorsal surface of urohyal keel, with posterior tip situated immediately in front of anteromedial region of contralateral hypobranchials 3. Ossification of basibranchial 2 nearly as long as bony portions of basibranchial 3 and hypobranchial 1. Basibranchial 4 completely cartilaginous, represented by a hexagonal plate. Basibranchial 4 bordered anteriorly by hypobranchials 3, laterally by cartilaginous heads of ceratobranchials 4, and posteriorly by cartilaginous heads of ceratobranchials 5. Hypobranchial 1 rod-like, with cartilage only in its proximal and distal tips, and without uncinate process on its anterodistal portion. Hypobranchial 2 elongate, approximately trapezoidal, mostly cartilaginous except for pointed, anterolaterally-oriented, process on its anterodistal region. Hypobranchial 3 trapezoidal, closely positioned relative to its counterpart, mostly cartilaginous except for pointed, anterolaterally-oriented, process on its anterodistal region. Hypobranchial 4 absent. Five slightly curved and mostly ossified ceratobranchials present; with cartilage present only at their extremities.Proximal portion of ceratobranchial 2 with concavity at its posterior margin, which is posteriorly limited by pointed, mesially directed, process.Proximal portion of ceratobranchial 3 with shallow concavity at its posterior margin. Ceratobranchial 5 expanded posteromedially to support patch of fine conical teeth dorsally, arranged in 2-4 rows, with the teeth of most mesial row distinctly larger; 20-26 teeth in smaller specimen (32.5 mm SL) and 27-30 teeth in larger specimen (43.9 mm SL). Five epibranchials, first 3 rod-like and largely ossified except at tips. Anterior margin of epibranchial 1 with noticeable elongate uncinate process. Anterior border of epibranchial 2 with blunt process. Posterior margin of epibranchial 3 with robust uncinate process. Epibranchial 4 with rectangular aspect, produced by broad anterior and posterior crests. Epibranchial 5 curved, completely cartilaginous, and situated medial to distal cartilaginous head of ceratobranchial 4, and reaching distal cartilaginous head of ceratobranchial 5. Pharyngobranchials 1 and 2 absent. Pharyngobranchial 3 elongate, rod-like, with shallow medial lamina, and with lateral margin smooth. Pharyngobranchial 4 well ossified and elongate. Neomorphic cartilaginous nodule, usually located near inner tips of epibranchials 1 and 2 and anterior tip of pharyngobranchial 3, absent. Large, elongate upper pharyngeal tooth plate situated immediately posterior to posterior extremity of pharyngobranchial 3, ventral to pharyngobranchial 4, and medial to tips of epibranchials 3 and 4. Upper pharyngeal tooth plate with mesial margin plicate, ventrally supporting patch of fine conical teeth, arranged in 2-4 rows, with 4-5 most posterior teeth of main row very large and curved at their tips; 27-28 teeth in smaller specimen (32.5 mm SL) and 29-34 teeth in larger specimen (43.9 mm SL).
Pectoral-fin rays I+6 (17*), rarely I+7 (1). Fin well-developed, rounded, and thick, with insertion immediately posterior to opercular membrane. Tip of first unbranched pectoral-fin ray prolonged (filamentous portion of ray 20-30% of total ray length in larger individuals and proportionally shorter in smaller individuals). Muscular base of pectoral fin broad, its breadth about 60% of mouth width.
Pelvic-fin rays I+4 (18*), distal border rounded. Insertion of pelvic fin slightly anterior of vertical through origin of dorsal fin. Inner margin of bases of pelvic fins very close to each other, sometimes in contact. Tip of adpressed pelvic fin extending beyond urogenital and anal openings, but falling short of origin of anal fin. Pelvic splint very thin and slightly shorter than first pelvic-fin ray.
Dorsal-fin rays ii+II+7 (17*), rarely iii+III+7 (1). Base of fin short with distal profile convex. Fin with 8 slender, free basal radials (medial and distal radials fused). Small bony piece fitted in posterior space of distal tip of last basal radial of dorsal fin. Origin of dorsal fin posterior to middle of total length. Tip of first basal radial of dorsal fin immediately posterior to neural spines of 22nd vertebra; tip of its last basal radial immediately anterior to neural spine of 27th vertebra.
Anal-fin rays ii+II+5 (9), iii+II+5 (9*). Base of fin short. Fin with 6-7 slender and free basal radials. Origin of anal fin slightly anterior to vertical through base of last dorsal fin-ray. Tip of first basal radial of anal fin immediately posterior to hemal spines of 26th vertebra, tip of its last basal radial immediately anterior to hemal spine of 30th-31st vertebrae.
Caudal-fin rays 45 (2), 46 (3), 47 (3), 48 (1), 49 (1), 50 (3). Fin wide, rounded distally, upper lobe slightly longer than lower lobe ( Figs. 1-3 View Fig View Fig View Fig ). Procurrent caudal-fin rays slightly more numerous dorsally, 23 (1), 24 (2), 25 (6), 26 (1), 27 (2), 28 (1), than ventrally, 20 (1), 21 (2), 22 (6), 23 (3), 24 (1). Upper caudalfin lobe with 5 branched rays (18*); lower caudal-fin lobe usually with 6 branched rays (17*), rarely 5 (1). Specimens 29.6 mm SL or smaller with branched caudal-fin rays splitting once; specimens 30.7 mm SL or larger typically with medial rays of caudal fin branched twice. Parhypural and hypurals 1 and 2 co-ossified and jointly bearing 7 rays. Hypural 3 autogenous, with 3 rays. Hypural 4 and 5 totally fused to each other, and jointly bearing 3 rays. Hypurapophysis and secondary hypurapophysis fused to form horizontal and shallow shelf extending to hypural 1, complex hypurapophysis (hypurapophysis “ type B” of Lundberg & Baskin, 1969). Epural and opistural cartilage absent.
Total vertebrae count 42. Pleural ribs 14-15. First pleural rib straight and much thicker than second rib. First complete hemal spine on 20th-21st vertebra.
Head sensory canals with simple (non-dendritic) tubes ending in single pores ( Figs. 4-5 View Fig View Fig ). Supraorbital sensory canal continuous and connected to optic and infraorbital sensory canals posteriorly. Supraorbital sensory canal bearing 3 branches and pores s1, s3, and s6 (epiphyseal branch and pore). S2, s4, s5, s7, and s8 (parietal) branches and pores absent. Otic sensory canal short, without pores, and continuous with posterior limits of supra- and infraorbital sensory canals, anteriorly, and with anterior limit of postotic sensory canal, posteriorly. Postotic (or temporal) sensory canal extends from posterior limit of otic sensory canals to anterior limit of lateral line, bearing 2 branches and pores (po1 and po2). Infraorbital sensory canal with 2 segments, anterior with 2 branches and pores (i1 and i3) and posterior segment with 2 branches and pores (i10 and i11). Preoperculomandibular sensory canal absent. Lateral-line canal very short, with 2 pores, extending from posteriormost region of opercle to above middle of pectoral fin. Conspicuous rostral line of neuromasts surrounding anterior limit of anterior nostril. Interrupted line of superficial neuromasts along midlateral portion of trunk, from above pectoral fin-base to caudal-fin base, visible in some specimens.
Color. Ground color of dorsal part of head and body pale yellow to orange yellow from mouth to base of tail, mottled with superimposed grayish brown to dark brown medium-sized blotches coalescing irregularly, interspersed with smaller light brown to grayish brown blotches and spots in life ( Fig. 3b View Fig ). Pigmentation in 2 skin layers. Lower layer composed of larger horizontally-elongate blotches formed of densely grouped chromatophores. Upper layer composed of smaller blotches and spots formed by loosely arranged chromatophores often overlaying larger blotches ( Fig. 4 View Fig ). Trunk with blotches of lower layer generally arranged in 4, irregular, longitudinal, rows (particularly evident in larger specimens): 1 along dorsal region composed of relatively scattered blotches; 1 along dorsolateral region, 1 along midlateral region (most conspicuous), and 1 along ventrolateral region. Each row formed of large, horizontallyelongate, well-defined blotches. Trunk pigmentation interrupted by thin unpigmented lines running along limits of myomeres, particularly visible on ventrolateral region of body. Ground color of head and lateral surface of body as described above, but with blotches and spots less numerous and concentrated than on dorsal portion of body. Barbels light yellow with light brown spots dorsally and whitish ventrally.Ventrolateral surface of head and belly white. Color pattern of juveniles grayish and lighter overall ( Fig. 3a View Fig ).
Dorsal and anal fins light yellow with light to dark brown spots concentrated on rays in life ( Figs. 1-4 View Fig View Fig View Fig View Fig ). Caudal fin light yellow to orange yellow with light to dark brown pigmentation concentrated on rays and arranged in interrupted narrow bands in most specimens in life. Dorsal surface of pectoral fin hyaline to light yellow with limited light brown pigmentation basally in smallest specimen in life; pigmentation spreading distally in larger individuals in life, especially along first ray. Pelvic fins hyaline. In alcohol yellow color fades to light tan.
The color pattern alone allows to distinguish the new species from the known species of Trichomycterus from southeastern Brazil outside the T. brasiliensis species-complex: T. concolor Costa has body color mostly uniform, subtly denser on the back and along the midlateral line; T. mirissumba Costa has laterodorsal, lateral, and lateroventral wide and ill-defined bands almost coalescent, composing an almost uniform pattern; T. reinhardti (Eigenmann) has blotches of laterodorsal, lateral, and ventrolateral rows arranged as longitudinal stripes, the lateral one broadest; T. paolence and T. triguttatus have irregularly distributed spots and/or dots, some of them arranged in three faint longitudinal rows; T. variegatus has blotches of deeper layer arranged in four ill-defined stripes, each of them sometimes interrupted and coalesced with the adjacent stripes; and T. zonatus (Eigenmann) has large, irregular blotches arranged in dorsal, laterodorsal, lateral, and ventrolateral rows, vertically coalescent in bars.
Etymology. The specific epithet “maracaya” (also “mbaracaya”) is a Tupi-Guarani Amerindian name for the margay wild cat ( Leopardus wiedii ), an allusion both to the blotched pigmentation pattern of the new species and its predatory habits on vertebrates (tadpoles). A noun in apposition. We propose the English common name Margay pencil catfish for this species.
Distribution. Known only from an unnamed streamlet running along the foothill of the Morro do Ferro, município de Poços de Caldas ( Fig. 9 View Fig ), State of Minas Gerais, southeastern Brazil [see Bonotto (1994) for figures of the Morro do Ferro area]. The streamlet is a tributary of the rio Pardo system (upper rio Paraná).
Natural history notes. Trichomycterus maracaya was found along a stretch of about 400 m of a streamlet 60-210 cm wide with maximum depth about 140 cm running through a semi-deciduous gallery forest ( Fig. 10 View Fig ). The streamlet runs through a yellowish limonite (iron hydroxide) rocky bed with pebbles and sand, and several pools with muddy bottom and accumulated plant litter and fine sediment. Small individuals (up to 32.5 mm SL) were found among submerged roots near the bank as well as buried in plant litter in small pools 30-50 cm deep.Larger individuals (up to 51.3 mm SL), in contrast, were found only among plant litter in larger pools up to 100 cm deep. Trichomycterus maracaya was the only fish species recorded in the streamlet, along with the tadpoles of three species of stream-dwelling frogs. The fishes were active at dawn and night, foraging on the exposed areas of the stream bed and on the leaf litter. One large individual (about 50 mm SL) was observed at early night swallowing tail-first a young, leaf litter-dwelling tadpole of the glass frog Hyalinobatrachium sp. (Centrolenidae) . Four other individuals (45.6-51.3 mm SL) regurgitated young tadpoles of the same frog species upon handling. The regurgitation was digested to differing degrees, an indication that the catfish may prey on tadpoles in the litter throughout the day. In addition to tadpoles, immature benthic aquatic insects (Ephemeroptera, Trichoptera, and Plecoptera) were found among the food regurgitated by these four individuals. Smaller individuals (22.8-33.6 mm TL) had fed on immature benthic aquatic insects, a diet recorded for most Trichomycterus species (e.g., Castro & Casatti, 1997; de Pinna, 1998; Ferriz, 1998; IS and FAB, pers. obs.).








| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |