
Monatractides polyhymnia, Pešić, Vladimir & Smit, Harry, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3820.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BBE4177B-5A2C-4911-987C-454BB8FA767C |
|
DOI |
https://doi.org/10.5281/zenodo.6143473 |
|
persistent identifier |
https://treatment.plazi.org/id/03BFDC60-AB70-6929-FF47-F946FCCFFC2C |
|
treatment provided by |
Plazi |
|
scientific name |
Monatractides polyhymnia |
| status |
sp. nov. |
Monatractides polyhymnia n. sp.
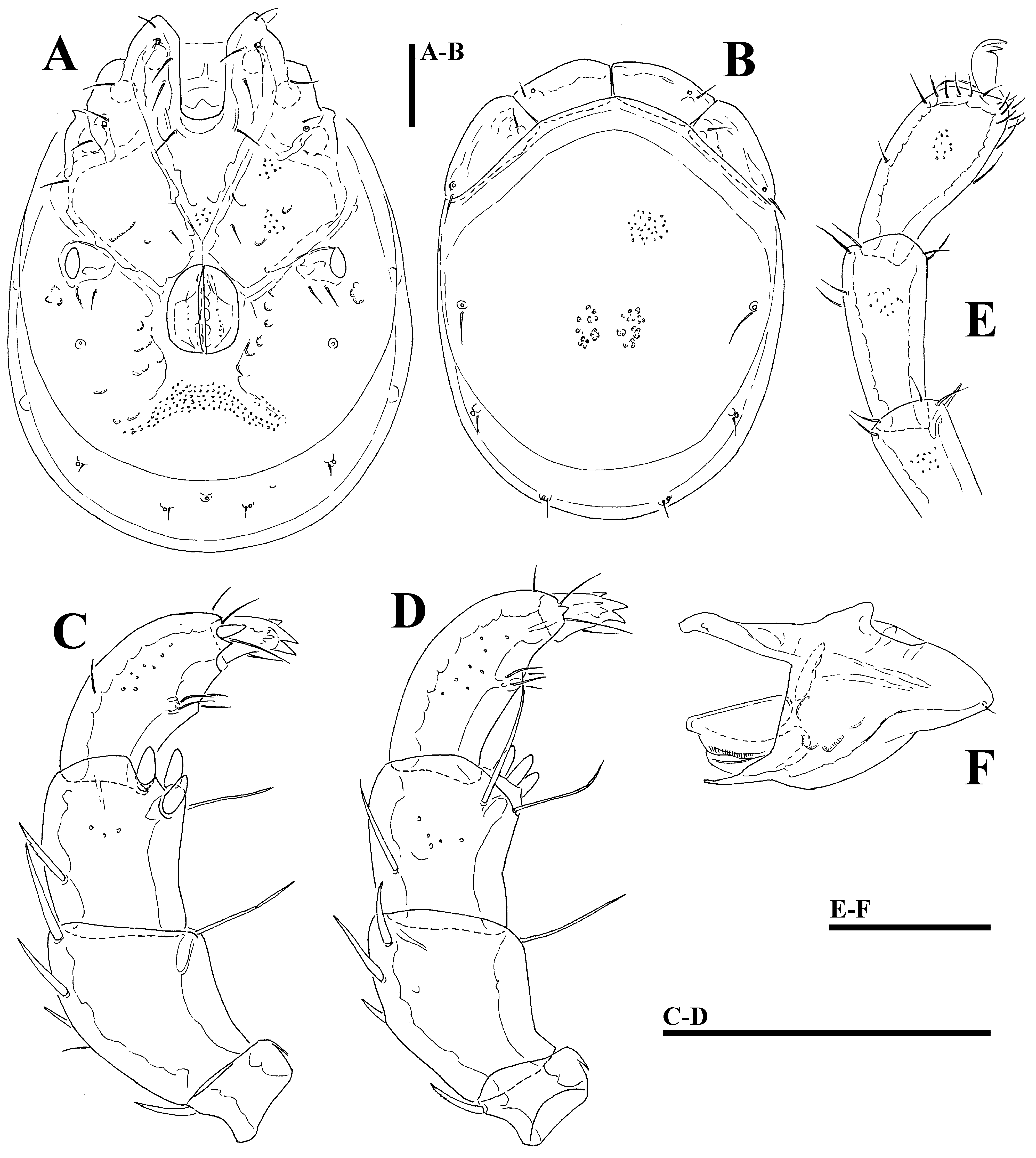
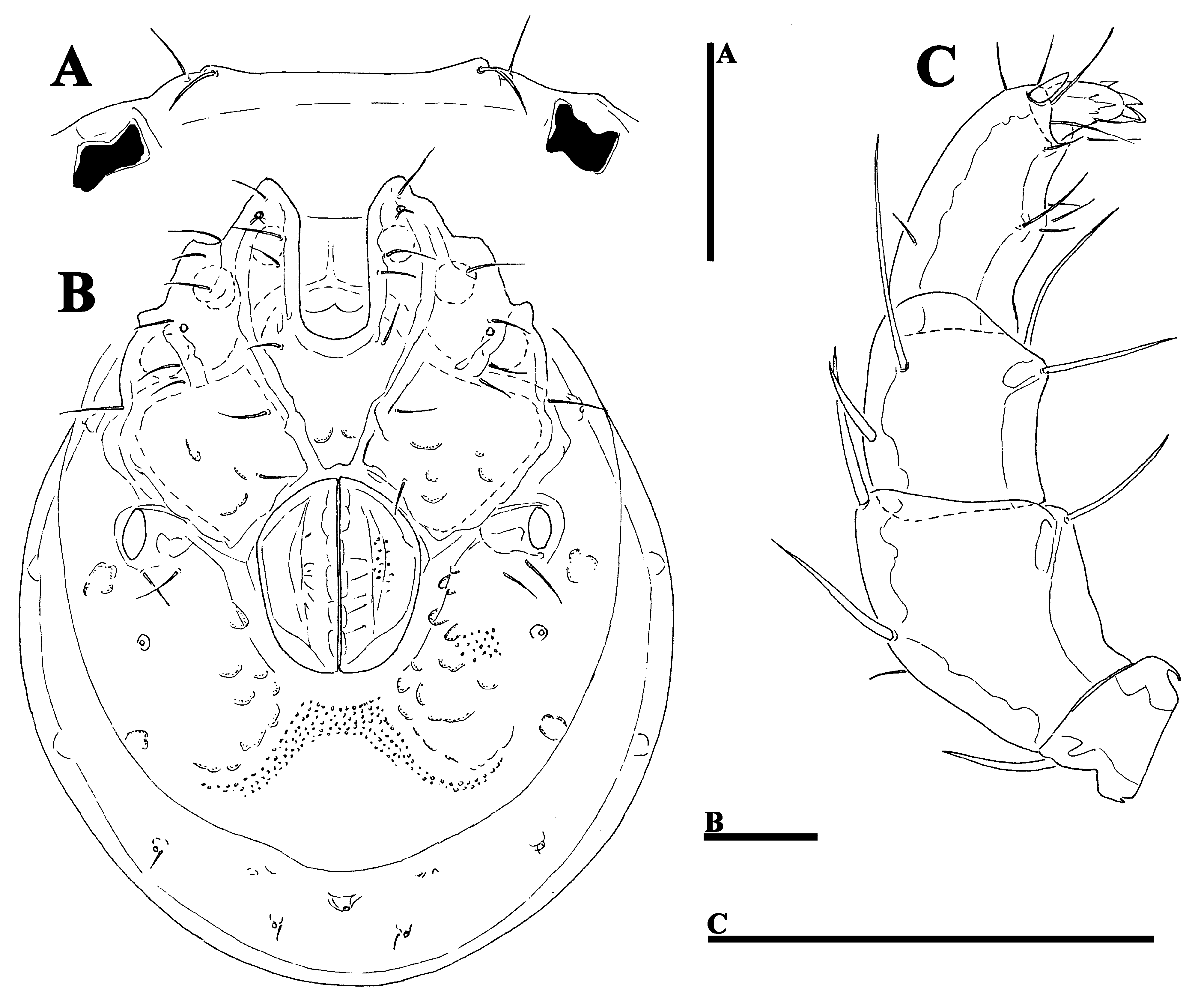
( Figs. 30A–F View FIGURE 30 A – F , 31A–C View FIGURE 31 A – C , 37E–F View FIGURE 37 A – I , 38E–F View FIGURE 38 A – I )
Type series. Holotype male, dissected and slide mounted, Ghana: Kue River, Kyabobo NP, 8º 31.087 N, 0º 36.049 E, alt. 208 m asl., 25.ii.2013, Smit. Paratypes: 1/1/0, same data as holotype, 0/1/0 dissected and slide mounted.
Diagnosis. P-3 ventral margin concave, sexual dimorphism of P-3, P-4 bowed, ventral margin with setaebearing protrusion in distal half of segment. Male: genital field oval (L/W ratio 1.26); P-3 with three stout, medially orientated setae at the ventrodistal margin. Female: genital field elongated trapezoid (L/W ratio 1.2); P-3 distally with ventrolateral seta.
Description. General features —Idiosoma roundish; frontal platelets shorter than shoulder platelets (shoulder/ frontal platelets L ratio 1.1–1.2); frontal margin with flat protrusions medial to the eyes ( Fig. 31A View FIGURE 31 A – C ); the tips of Cx-I orientated to the lateral side. Cxgl-4 located far anteriorly, near insertion of I-L; gnathosomal bay deep U-shaped, proximally wide box-shaped; posterior suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field and directed posterolaterally; excretory pore and Vgl–2 away from the line of primary sclerotization, Vgl–2 posterior to excretory pore; gnathosoma nearly triangular in lateral view, with short, cone-shaped rostrum ( Fig. 30F View FIGURE 30 A – F ); palp compact, ventral margin of P-2 proximally with sharp bend, distally straight, bearing ventral seta, P-3 ventral margin concave, sexual dimorphism of P-3, P-4 bowed, ventral margin with setae-bearing protrusion in distal half of segment. Male: medial margin of Cx-II/III relatively short; genital field small and oval in shape; P-3 ventrodistally bearing three stout (short and thick) setae, orientated medially and one ventrolateral seta ( Figs. 30C–D View FIGURE 30 A – F ). Female: suture lines Cx-I/II complete, not meeting in the medial line, but separately ending at anterior genital field margin ( Fig. 31B View FIGURE 31 A – C ); genital field pentagonal; P-3 distally with ventrolateral seta ( Fig. 31C View FIGURE 31 A – C ).
Measurements. Male (holotype)—Idiosoma (ventral view: Figs. 30A View FIGURE 30 A – F , 38E View FIGURE 38 A – I ) L 665, W 499; dorsal shield ( Figs. 30B View FIGURE 30 A – F , 37E View FIGURE 37 A – I ) L 559, W 418, L/W ratio 1.34; dorsal plate L 516; shoulder platelets L 142–147, W 69–70, L/W ratio 2.07–2.09; frontal platelets L 120–127, W 58–59, L/W ratio 2.03–2.2; shoulder/frontal platelets L ratio 1.12–1.22. Gnathosomal bay L 131, Cx-I total L 281, Cx-I mL 150, Cx-II+III mL 22; ratio Cx-I L/Cx-II+III mL 12.8; Cx-I mL/Cx-II+III mL 6.8. Genital field L/W 106/84, ratio 1.26; distance genital field-excretory pore 175, genital fieldcaudal idiosoma margin 241. Gnathosoma vL 184; palp total L 232–233, dL/H, dL/H ratio: P-1, 21–22/28, 0.78; P- 2, 68/47, 1.43; P-3, 49/43, 1.15; P-4, 68/28, 2.4; P-5, 26/13, 2.0; P-2/P-4 ratio 1.0; dL of I-L-5–6 (Fig.): 122, 128; I-L-6 H 58–59, dL/H I-L-6 ratio 2.2.
Female (paratype from Kue River, n = 1)—Idiosoma (ventral view: Figs. 31B View FIGURE 31 A – C , 38F View FIGURE 38 A – I ) L 731, W 584; dorsal shield ( Fig. 37F View FIGURE 37 A – I ) L 641, W 494, L/W ratio 1.3; dorsal plate L 594; shoulder platelets L 156–159, W 78–81, L/W ratio 2.0; frontal platelets L 131–134, W 67, L/W ratio 1.95–2.0; shoulder/frontal platelets L ratio 1.19. Gnathosomal bay L 147, Cx-I total L 267. Genital field L/W 173/145, ratio 1.19; distance genital field-excretory pore 209, genital field-caudal idiosoma margin 284. Gnathosoma vL 199; chelicera total L 240; palp total L 229, dL/H, dL/H ratio: P-1, 23/28–29, 0.81; P-2, 69/48, 1.43; P-3, 48/38–39, 1.24; P-4, 64/29, 2.16; P-5, 25/12, 2.06; P- 2/P-4 ratio 1.09; dL of I-L-2–6: 69, 92, 111, 122, 131; I-L-6 H 55, dL/H I-L-6 ratio 2.4.
Etymology. The species is named after Polyhymnia (Ancient Greek: Πολυύμνια), one of nine Muses from Greek mythology, who was a patron of sacred poetry and song. The species name is a noun in apposition (in the nominative case).
Discussion. The new species shows a remarkable sexual dimorphism, as in the male P-3 has a group of three stout, medially orientated setae at the ventrodistal margin, while these setae are lacking in the female. Due to the shape of P-3 with stout, medially oriented setae at the ventrodistal margin, the new species from Ghana seems to be closely related to M. spinipalpis ( K.O. Viets, 1981) a species described by K.O. Viets (1981) based on a single male from South Africa. Later on, Goldschmidt & Smit (2009) reported a single male of M. spinipalpis from Kwazulu- Natal Province and provided an additional description of this species. In the original description, K.O. Viets (1981) described this species as close to M. mitchelli ( K.O. Viets, 1965) , but differing in a stout, medially oriented setae at the ventrodistal margin of P-3 lacking in the latter species. Obviously, he was not aware that the difference in the shape of P-3 might be results of the sexual dimorphism, as the latter species was known only from two females from Transvaal ( K.O. Viets 1965). It is possible that M. mitchelli represents a senior synonym of M. spinipalpis but this should be checked by finding additional material, including the male, from the locus typicus of M. mitchelli .
Monatractides spinipalpis differs from the new species from Ghana in larger idiosoma dimensions, a more elongated genital field (L/W ratio 1.41–1.55, data taken from K.O. Viets 1981 and Goldschmidt & Smit 2009), P-2 shorter than P-4 and by having a well developed ventrodistal protrusion on P-3. The female of M. mitchelli can be separated by the more elongated genital field, strongly tapering to the caudal end (see K.O.Viets 1958, Fig. 9 View FIGURE 9 A – C ).
Remarks. No ejaculatory complex of Monatractides spinipalpis has been described yet (see: K.O. Viets 1981, Goldschmidt & Smit 2009) Also in males in the present study this organ was not found, but sclerotized framework immediately above the genital field can be hardly visible.
Habitat. A sandy/bouldary streams, shaded by riparian vegetation (see Fig. 45 View FIGURE 45 C in Pešić et al. 2013).
Distribution. Ghana; known only from the type locality.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |