
Bryconamericus misei, Pedroso & Deprá & Pavanelli, 2024
|
publication ID |
https://doi.org/ 10.1590/1982-0224-2023-0049 |
|
publication LSID |
lsid:zoobank.org:pub:52E17D1B-5F1B-4A06-AD00-2CF469464C61 |
|
DOI |
https://doi.org/10.5281/zenodo.11126694 |
|
persistent identifier |
https://treatment.plazi.org/id/03BF87DC-FFDA-FF8B-31F7-BB48EED8F8ED |
|
treatment provided by |
Felipe |
|
scientific name |
Bryconamericus misei |
| status |
sp. nov. |
Bryconamericus misei , new species
urn:lsid:zoobank.org:act:2CAF55C7-5B39-4C05-9318-8C69B783565D
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Tab. 1 View TABLE 1 )
Bryconamericus aff. iheringii View in CoL (non Boulenger, 1887). — Cavalli et al., 2018:5 (checklist of species of Piquiri River; citation).
Holotype. NUP 24255 , 54.8 mm SL, male, Brazil, Paraná State, Campina do Simão, unnamed stream, tributary to the rio Capivara , rio Piquiri basin, 25°05’43.02”S 51°49’29.00” W, 14 Jun 2019, F. T. Mise. GoogleMaps
Paratypes. All from the Paraná State, upper rio Paraná basin, Brazil. MZUEL 18401 , 5, 42.3–46.0 mm SL, Ortigueira, rio Apucarana , tributary to the rio Tibagi , 24°05’26.82”S 51°00’54.66”W, 21 Sep 2017, I. Vitture, A. Souza, R. Ono & N. Narezzi. GoogleMaps MZUEL 18438 , 3, 26.4–42.4 mm SL, córrego Água Boa , tributary to the rio Apucarana , tributary to the rio Tibagi , 23°57’40.43”S 51°01’17.46”W, 14 Nov 2017, I. Vitture, E. Santana, A. Souza & R. Ono. GoogleMaps NUP 15777, 5, 41.9–53.7 mm SL, Goioxim , rio Bonito , tributary to the rio Piquiri , 25°04’25.80”S 52°04’05.79”W, 26 Jan 2014, W. J. Graça, W. M. Domingues, F. A. Teixeira & R. J. Graça. GoogleMaps NUP 16042 , 3, 36.3–45.0 mm SL, Luiziana, rio Laranjeiras , tributary to the rio Ivaí , 24°20’26.85”S 52°11’42.50”W, 28 Jan 2014, W. J. Graça, W. M. Domingues, F. A. Teixeira & R. J. Graça. GoogleMaps NUP 18274 , 11, 35.9–47.8 mm SL, same locality as the holotype, 3 Oct 2015, W. J. Graça, F. A. Teixeira, W. M. Domingues & A. Frota. GoogleMaps NUP 18277 , 9, 27.0– 38.2 mm SL, Campina do Simão , unnamed stream, tributary to the rio Capivara , rio Piquiri basin, 25°08’38.49”S 51°49’29.01”W, 3 Oct 2015, W. J. Graça, F. A. Teixeira, W. M. Domingues & A. Frota. GoogleMaps NUP 24149 , 3, 18.4–54.9 mm SL, same locality as the holotype, 7 Dec 2018, F. T. Mise. GoogleMaps NUP 24150 , 8, 25.8–49.8 mm SL, same locality as the holotype, 14 Jun 2019, F. T. Mise. GoogleMaps NUP 24151 , 3, 44.0– 56.9 mm SL, same locality as the holotype, 16 Oct 2018, F. T. Mise. GoogleMaps NUP 24152 , 1, 39.1 mm SL, Campina do Simão , unnamed stream, tributary to the rio Capivara , rio Piquiri basin , 25°08’38.49”S 51°49’29.01”W, 22 Mar 2019, F. T. Mise. GoogleMaps NUP 24153 , 2, 37.8–50.2 mm SL, same locality as NUP 24152, 30 Nov 2019, F. T. Mise. GoogleMaps NUP 24154 , 7, 36.5–45.4 mm SL, same locality as the holotype, 30 Nov 2019, F. T. Mise. GoogleMaps NUP 24155, 18, 20.6–44.7 mm SL, same locality as NUP 24152, 14 Jun 2019, F. T. Mise GoogleMaps .
Diagnosis. Among the Stevardiinae genera endemic to the Southern Neotropics, Bryconamericus misei can be distinguished from species of Glandulocaudini, Bryconamericus sensu stricto (i.e., B. exodon and B. stramineus Eigenmann, 1908 ; Mirande, 2019), some species of Diapoma Cope, 1894 , Hysteronotus Eigenmann, 1911 , Lepidocharax Ferreira, Menezes & Quagio-Grassiotto, 2011 , Piabarchus Myers, 1928 , Piabina Reinhardt, 1867 , Planaltina Böhlke, 1954 , and Pseudocorynopoma Perugia, 1891 by having only a few, unmodified scales on the very base of the caudal fin (vs. presence of modified scales associated with glandular tissue on the ventral caudal-fin lobe of males and females of Planaltina and Diapoma pyrrhopteryx Menezes & Weitzman, 2011 , D. speculiferum Cope, 1894 , D. terofali (Géry, 1964) , and D. thauma Menezes & Weitzman, 2011 and of males of Hysteronotus , and Pseudocorynopoma ; presence of modified scales associated with glandular tissue on the dorsal caudal-fin lobe of males of Glandulocaudini; and non-modified scales covering more than one third of each caudal-fin lobe in Lepidocharax [also Knodus ]); anal-fin origin approximately at a vertical through the dorsal-fin terminus (vs. anterior to a vertical through the middle of the dorsal-fin base in Glandulocaudini [except Lophiobrycon Castro, Ribeiro, Benine & Melo, 2003 ], Hysteronotus , Lepidocharax , Piabarchus , Planaltina , and Pseudocorynopoma ); 8 total pelvic-fin rays (a single specimen with 7; vs. 7 in Diapoma , Lepidocharax , and Planaltina ); retrognathous mouth (vs. isognathous or prognathous in Glandulocaudini, Diapoma , Hysteronotus , Lepidocharax , Planaltina , and Pseudocorynopoma ; and teeth in the outer premaxillary series arranged in a regular line (vs. not aligned in Bryconamericus sensu stricto and Piabina [also in Creagrutus Günther, 1864 and ‘ B. ’ coeruleus Jerep & Shibatta, 2017, ‘ B. ’ mennii Miquelarena, Protogino, Filiberto & López, 2002 , ‘ B. ’ turiuba Langeani, Lucena, Pedrini & Tarelho-Pereira, 2005 , and Knodus moenkhausii (Eigenmann & Kennedy, 1903) ]).
Another lineage of Stevardiinae endemic to Southern Neotropics includes the nominal genera Hypobrycon Malabarba & Malabarba, 1994 , Nantis Mirande, Aguilera & Azpelicueta, 2006 and Odontostoechus Gomes, 1947 , in addition to ‘ B. ’ iheringii and several similar species currently assigned to ‘ Bryconamericus ’ ( Mirande, 2019; B. misei appears to belong in that lineage). Bryconamericus misei is distinguished from all members of that lineage, except for ‘ B.’ ecai da Silva, 2004 , ‘ B. ’ eigenmanni (Evermann & Kendall, 1906) , ‘ B. ’ ikaa Casciotta, Almirón & Azpelicueta, 2004 , ‘ B. ’ sylvicola Braga, 1998 and ‘ B. ’ ytu Almirón, Azpelicueta & Casciotta, 2004 by having a vertically elongated black humeral spot, followed by a second inconspicuous spot vs. a single vertically extended spot in Nantis indefessus Mirande, Aguilera & Azpelicueta, 2004 , Hypobrycon , ‘ B. ’ agna Azpelicueta & Almirón, 2001 , ‘ B. ’ microcephalus (Miranda Ribeiro, 1908) , ‘ B. ’ ornaticeps Bizerril & Perez-Neto, 1995 , ‘ B. ’ patriciae da Silva, 2004 , ‘ B. ’ rubropictus ( Berg, 1901) , and ‘ B. ’ tenuis Bizerril & Auraujo, 1992 ; a single, rounded humeral spot in Odontostoechus lethostigmus , ‘ B. ’ lambari Malabarba & Kindel, 1995 , ‘ B. ’ pyahu Azpelicueta, Casciotta & Almirón, 2003 and ‘ B. ’ uporas Casciotta, Azpelicueta & Almirón, 2002 ; Bryconamericus misei differs from ‘ B. ’ ecai , ‘ B. ’ eigenmanni , ‘ B. ’ iheringii , ‘ B. ’ aff. iheringii from the upper rio Paraná ( Frota et al., 2016; Reis et al., 2020), ‘ B. ’ ikaa , ‘ B. ’ sylvicola and ‘ B. ’ ytu , as well from ‘ B. ’ microcephalus by having a body depth of 27.8–31.8% SL vs. 33.1–36.9% in ‘ B. ’ ecai , 33.7–42.3% in ‘ B. ’ iheringii , 34.2–39.3% in ‘ B. ’ aff. iheringii ( Tab. 2 View TABLE 2 ), 33.7–36.4% in ‘ B. ’ ikaa , 23.5–27.7% in ‘ B. ’ microcephalus , 36.1–40.7% in ‘ B. ’ sylvicola and 34.6–37.9% in ‘ B. ’ ytu .
In addition, Bryconamericus misei differs from ‘ B. ’ eigenmanni , and ‘ B. ’ iheringii by having 13–16 total external gill rakers on first branchial arch vs. 17–21; from ‘ B. ’ pyahu by having 6–7 gill rakers on epibranchial vs. 4–5 in ‘ B. ’ pyahu ; from adults of Odontostoechus lethostigmus by having two rows of teeth on the premaxilla vs. a single row; from Nantis indefessus by having four teeth in the inner row of premaxilla vs. five; from ‘ B. ’ ornaticeps and ‘ B. ’ sylvicola by having 16–19 branched anal-fin rays vs. 14–15 in ‘ B. ’ ornaticeps and 22–25 in ‘ B. ’ sylvicola ; from ‘ B. ’ agna and ‘ B. ’ uporas by having 3–5 cusps on the innerseries premaxillary teeth vs. 7; from ‘ B. ’ tenuis , by having distally compressed teeth vs. massive teeth; from Hypobrycon , by having dentary teeth positioned anterodorsally vs. teeth positioned along the anterior margin of dentary (compare Fig. 2D View FIGURE 2 with fig. 1 in Malabarba, Malabarba, 1994); from ‘ B. ’ agna and H. poi Almirón, Casciotta, Azpelicueta & Cione, 2001 by having 8–11 teeth on dentary vs. 6–7; from ‘ B. ’ microcephalus , ‘ B. ’ iheringii and from ‘ B. ’ aff. iheringii from the upper rio Paraná by having a lower orbital diameter (28.1–33.9% HL vs. 38.4–41.6% in ‘ B. ’ microcephalus , 33.6–39.7% in ‘ B. ’ iheringii and 34.8–40.9% in ‘ B. ’ aff. iheringii ; Tab. 2 View TABLE 2 ).
Description. Morphometric data presented in Tab. 1 View TABLE 1 . Dorsal profile of head convex from tip of snout to vertical through anterior border of nostrils, slightly convex from nostrils to dorsal-fin origin; straight from that point to adipose-fin origin, slightly concave along caudal peduncle. Ventral profile convex from tip of dentary to anal-fin origin; straight from that point to end of anal-fin, slightly concave along caudal peduncle.
Mouth slightly retrognathous, always positioned at level of ventral border of orbit or below ( Fig. 3 View FIGURE 3 ). Posterior tip of maxilla exceeding vertical through anterior limit of orbit. Outer premaxillary tooth row with three(3), four*(17) or five(10) tricuspid teeth; inner row with four*(30) tri to pentacuspid teeth; maxilla with two(2), three(12), four(14), five*(1) or six(1) tricuspid teeth; dentary with four large tri to tetracuspid teeth anteriorly, and four(7), five(17), six(2) or seven*(2) smaller teeth gradually decreasing in size posteriorly. External gill rakers on first arch 6*(13) or 7(17) on upper limb and 7(3), 8(17) or 9*(10) on lower limb, counted in entire specimens.
Scales cycloid. Lateral line completely pored, with 36(3), 37(13), 38*(11) or 39(1) perforated scales. Longitudinal scale rows between lateral line and dorsal-fin origin 4½(1), 5(2), 5½*(27) or 6½(1); longitudinal scale rows between lateral line and pelvic-fin origin 4(3), 5*(23) or 6(6); single row of 4(1), 5(4), 6(6), 7(8), 8(5) or 9*(5) scales on base of anteriormost anal-fin rays; circumpeduncular scales 14*(30); axillary scale present on pelvic-fin insertion.
Pectoral-fin rays I,9(1), i,10(1), i,10,i(11), i,11(2), i,11,i*(10), i,12(2) or i,12,i(3); tip of pectoral-fin not reaching pelvic-fin origin; pelvic-fin rays i,5,i(1), i,6,i(24) or i,7*(5); dorsal-fin rays ii,8*(30); first unbranched ray about one-half length of second unbranched ray; first branched ray longer than second unbranched ray; distal margin of dorsal-fin slightly rounded; anal-fin rays iii,16(3), 17*(19), 18(7) or 19(1); anal-fin insertion at vertical through insertion of last dorsal-fin ray or posterior to it; adipose-fin present, its insertion posterior to or at vertical through insertion of last anal-fin ray; caudal-fin principal rays i,17,i*(30); caudal-fin lobes rounded, equally sized.
Dorsal procurrent caudal-fin rays 12(2); ventral procurrent caudal-fin rays 11(2); total vertebrae 37(2); abdominal vertebrae 18(2); caudal vertebrae 19(2); fifth to seventeenth vertebrae type A; eighteenth vertebra type B; nineteenth to thirty-seventh vertebra type D; ribs 13(2), on fifth through seventeenth vertebra; supraneurals 5(2), between fourth and ninth vertebra; dorsal-fin pterygiophores 9(2), between eleventh through nineteenth vertebra; anal-fin pterygiophores 17(2), between nineteenth through twenty-seventh vertebra; two rows of gill rakers in arches 1–4, one row in fifth arch (only external); external rakers on first arch 17 (0, 1, 6, 0, 8, 0, 2); internal rakers on first arch 10 (0, 0, 6, 0, 4, 0, 0); external rakers on second arch 15 (0, 1, 5, 1, 6, 0, 2); internal rakers on second arch 12 (0, 0, 6, 0, 6, 0, 0); external rakers on third arch 14 (0, 1, 5, 1, 6, 0, 1); internal rakers on third arch 13 (0, 0, 5, 0, 8, 0, 0); external rakers on fourth arch 14 (0, 0, 6, 0, 8, 0, 0); internal rakers on fourth arch 7 (0, 0, 0, 0, 7, 0, 0); rakers on fifth arch 8 (only external).
Coloration in alcohol. Ground color yellow to dark-brown ( Fig. 1 View FIGURE 1 ). Dorsal portion of head with dark brown coloration from tip of snout to posterior margin of supraoccipital, extending posteriorly as dorsal band to end of caudal peduncle; great concentration of melanophores around orbit and along posterior margin of maxilla; fewer melanophores scattered on infraorbitals, interopercle and opercle, and anterior portion of lower jaw; melanophores also on distal margin of scales, more concentrated on scales above lateral line, forming reticulated pattern; vertically elongated black humeral spot across second to fourth lateral line scale, reaching three scale rows above and one scale row below lateral line, tapering downward. Second, inconspicuous humeral spot separated from first spot by two scales; dark midlateral stripe from second humeral spot to end of caudal peduncle; caudal-fin with narrower stripe along median rays and melanophores concentrated on middle portion of lobes; dorsal and anal fins with melanophores concentrated on distal border; first unbranched dorsal-fin ray completely covered by melanophores; pectoral, pelvic and adipose fins with few scattered melanophores.
Sexual dimorphism. Pelvic-fin slightly longer and more rounded in males ( Tab. 1 View TABLE 1 ), covering completely urogenital opening in ventral view, and usually reaching anal-fin origin; pelvic-fin slightly shorter and more pointed in females, not covering urogenital opening in ventral view ( Figs. 4C–D View FIGURE 4 ). Distal border of anal-fin straight in males, slightly concave in females ( Figs. 4A–B View FIGURE 4 ). Anal-fin base slightly longer in males, pre-anal distance slightly longer in females ( Tab. 1 View TABLE 1 ). Sexually mature males with bony hooks on anal and pelvic fins; anal-fin hooks on last unbranched ray through fifth to ninth branched ray; pelvic-fin hooks on all rays except first; bony hooks absent in females. Sexually mature males with breeding tubercles along dorsal and lateral portion of head, and on distal border of scales ( Fig. 3A View FIGURE 3 ). Tubercles absent in females or, if present, few and concentrated only on distal border of scales.
Geographical distribution. Bryconamericus misei is known from the rio Bonito and from two small tributaries to the rio Capivara, in the rio Piquiri basin; from the rio Laranjeiras, a tributary to the rio Formoso, in the rio Ivaí basin; and from the rio Apucarana, a tributary to the rio Tibagi ( Fig. 5 View FIGURE 5 ).
Ecological notes. At the sampling localities of Bryconamericus misei , the two tributaries to the rio Capivara, rio Piquiri basin ( Figs. 6A–B View FIGURE 6 ) are about 1–2 m wide and 0.2 m deep, and lie about 800–900 m a.s.l. One of these ( Fig. 6A View FIGURE 6 ) is the type-locality. The rio Laranjeiras, rio Ivaí basin ( Fig. 6C View FIGURE 6 ) is about 4 m wide and 0.3 m deep, and lies about 498 m a.s.l. at the sampling location. The bottom of both streams is composed of small stones and pebbles, and some stretches with sandy bottom. The vegetation is mainly shrubs with some grasses, and stretches of the river are protected by canopies of small trees.
Etymology. The specific name misei is a patronymic, given in honor of Fábio Teruo Mise, for his contributions to the ichthyological education of THP and for collecting part of the type-specimens of Bryconamericus misei . A noun in a genitive case.
Conservation status. Bryconamericus misei is known from tributaries to the Piquiri, Ivaí and Tibagi rivers and was collected from six different sites distributed along these basins. No threats to this species have been detected, therefore it has been classified as Least Concern ( LC) according to the International Union for Conservation of Nature ( IUCN) criteria and categories ( IUCN Standards and Petitions Committee, 2022).

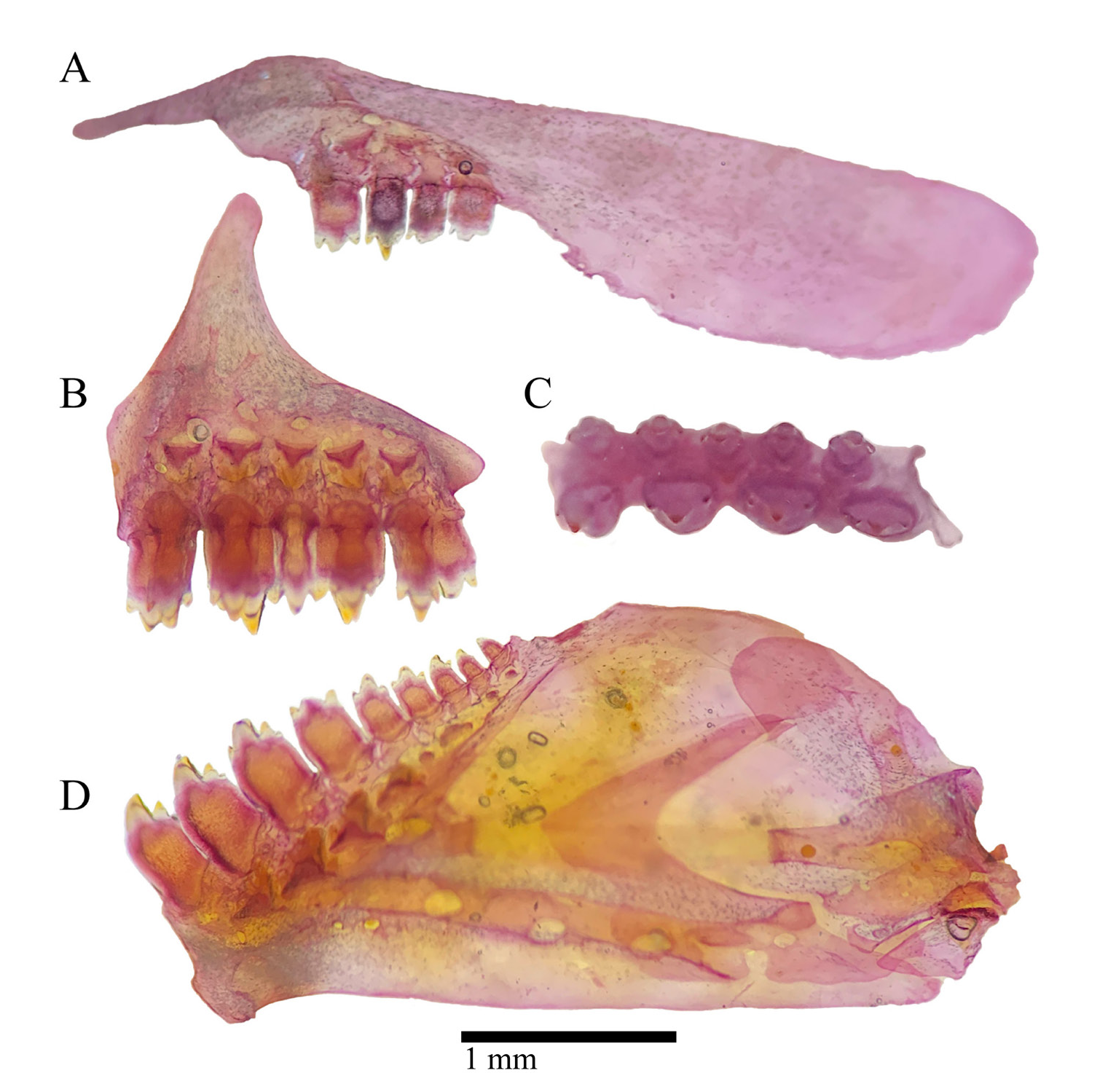

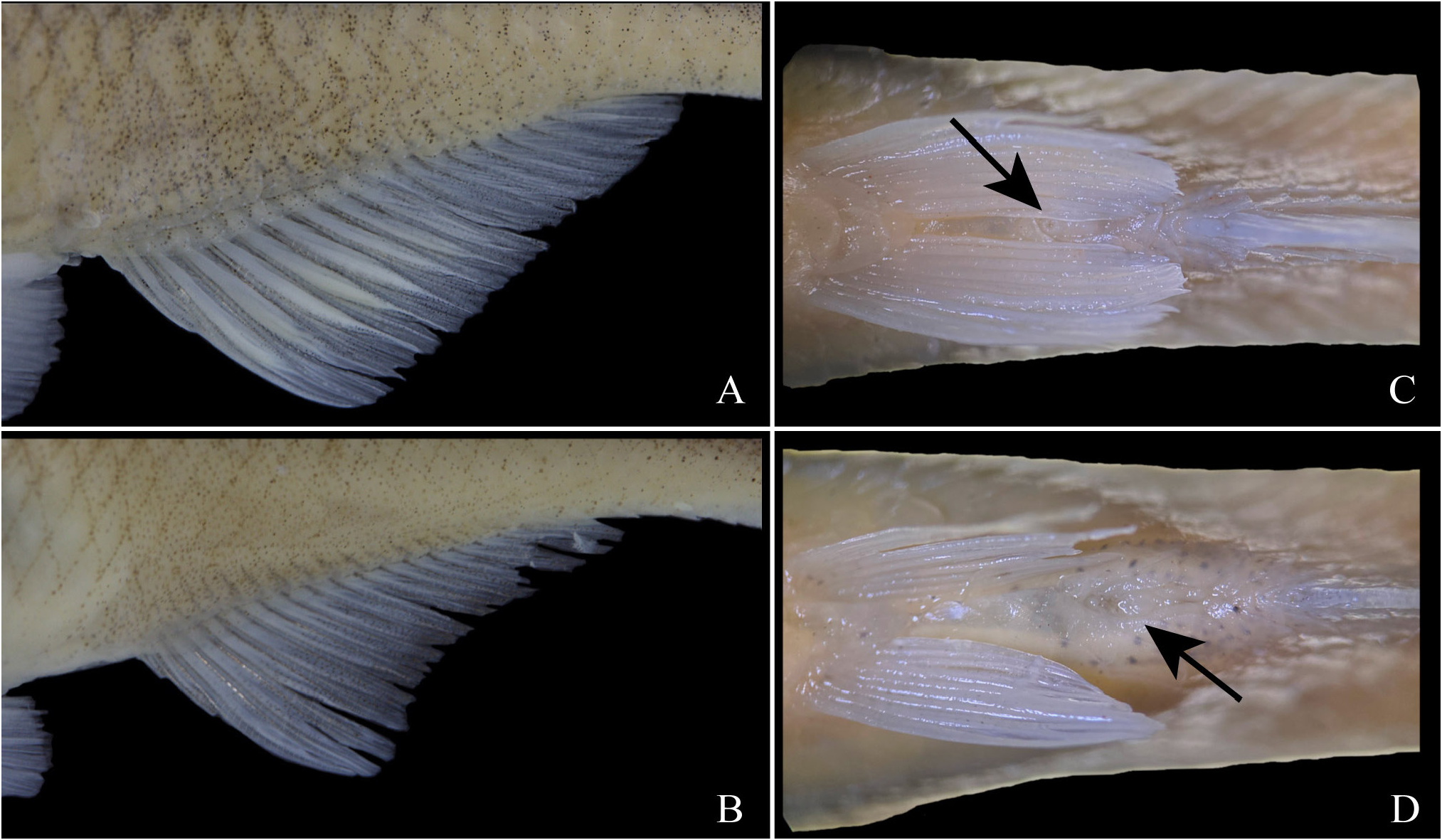
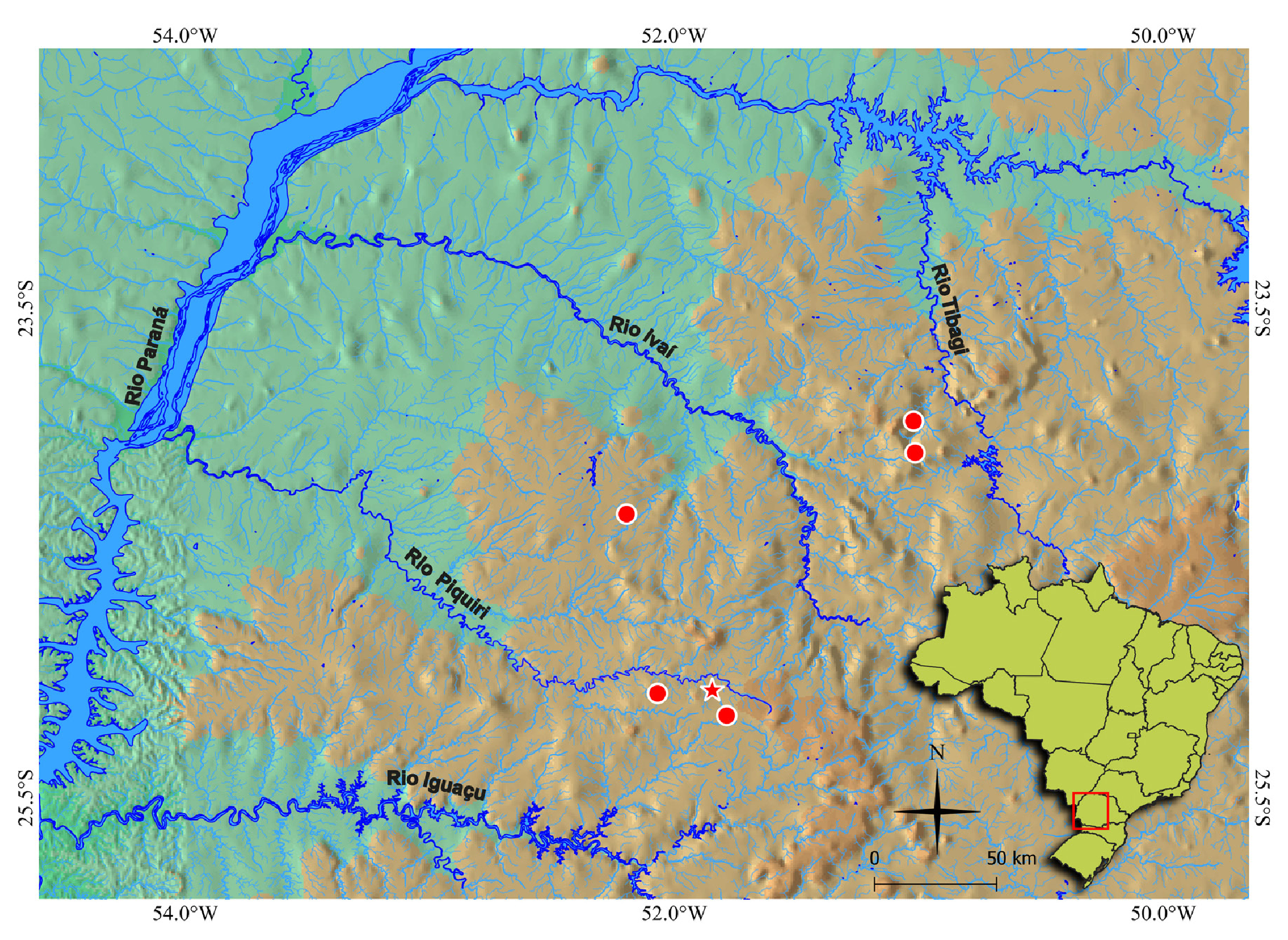

TABLE 1 | Morphometric data ofBryconamericus misei. N = number of specimens; SD = Standard deviation.
| Males | Females | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Characters | Holotype | N | Range | Mean | SD | N | Range | Mean | SD |
| Standard length (mm) | 54.8 | 16 | 39.1–50.2 | 43.4 | 13 | 40.7–56.9 | 46.5 | ||
| Percents of standard length | |||||||||
| Depth at dorsal fin origin | 29.4 | 16 | 28.9–31.8 | 30.3 | 0.7 | 13 | 27.8–31.7 | 30.0 | 1.0 |
| Snout to dorsal fin origin | 53.1 | 16 | 52.7–56.0 | 54.0 | 1.0 | 13 | 53.0–56.5 | 54.7 | 1.0 |
| Snout to pelvic fin origin | 46.9 | 16 | 45.0–48.6 | 47.0 | 1.1 | 13 | 46.5–49.0 | 47.7 | 0.8 |
| Snout to anal fin origin | 63.5 | 16 | 61.9–66.1 | 63.8 | 1.3 | 13 | 63.8–67.3 | 65.4 | 1.1 |
| Caudal peduncle depth | 12.4 | 16 | 11.1–13.3 | 12.2 | 0.5 | 13 | 11.1–12.2 | 11.7 | 0.3 |
| Caudal peduncle length | 15.0 | 16 | 14.3–17.3 | 15.8 | 0.9 | 13 | 14.3–17.1 | 15.5 | 0.7 |
| Pectoral fin length | 21.7 | 16 | 20.0–22.9 | 21.5 | 0.7 | 13 | 18.8–21.3 | 20.2 | 0.9 |
| Pelvic fin length | 15.5 | 16 | 14.4–17.7 | 16.5 | 0.8 | 13 | 13.9–15.7 | 14.8 | 0.6 |
| Dorsal fin length | 21.7 | 16 | 20.9–25.9 | 23.5 | 1.2 | 13 | 20.2–23.0 | 22.1 | 0.9 |
| Dorsal fin base length | 13.0 | 16 | 11.5–14.5 | 13.1 | 0.7 | 13 | 12.3–13.9 | 12.8 | 0.4 |
| Anal fin length | 17.3 | 16 | 16.9–20.5 | 18.6 | 1.0 | 13 | 15.8–18.8 | 17.2 | 0.7 |
| Anal fin base length | 25.4 | 16 | 21.2–26.6 | 24.6 | 1.3 | 13 | 20.8–23.9 | 22.6 | 0.9 |
| Dorsal fin origin to caudal fin base | 55.3 | 16 | 49.1–53.2 | 51.1 | 1.3 | 13 | 48.5–54.3 | 50.7 | 2.0 |
| Orbit to dorsal fin origin | 40.5 | 16 | 39.1–42.6 | 40.7 | 0.9 | 13 | 40.1–43.3 | 41.9 | 0.8 |
| Head length | 26.6 | 16 | 25.6–28.0 | 26.7 | 0.6 | 13 | 25.2–28.3 | 26.3 | 1.0 |
| Percents of head length | |||||||||
| Orbit diameter | 29.5 | 16 | 29.7–33.3 | 31.5 | 1.0 | 13 | 28.1–33.9 | 30.7 | 1.6 |
| Snout length | 24.7 | 16 | 22.9–25.7 | 24.2 | 0.8 | 13 | 22.0–27.8 | 25.4 | 1.5 |
| Interorbital width | 31.5 | 16 | 30.4–34.3 | 31.8 | 1.1 | 13 | 30.7–33.6 | 31.7 | 0.9 |
| Upper jaw length | 38.4 | 16 | 35.4–41.6 | 38.3 | 1.4 | 13 | 36.5–40.2 | 38.6 | 1.3 |
TABLE 2 | Morphometric data of ‘Bryconamericus’ iheringii from the Laguna dos Patos in comparison with ‘B.’ aff. iheringii from the upper rio Paraná basin. The diagnostic characters between these two species and B. misei are highlighted in bold. N = number of specimens.
| ‘ B.’ iheringii Laguna dos Patos | ‘ B. ’ iheringii upper rio Paraná | |||||
|---|---|---|---|---|---|---|
| Character s | ||||||
| Range | N | Mean | Range | N | Mean | |
| Standard length (mm) | 43.3–61.2 | 30 | 50.6 | 44.2–58.2 | 20 | 51.0 |
| Percents of standard length | ||||||
| Depth at dorsal fin origin | 33.7–42.3 | 30 | 36.2 | 34.2–39.3 | 20 | 36.9 |
| Snout to dorsal fin origin | 52.3–58.1 | 30 | 55.7 | 53.0–57.0 | 20 | 55.4 |
| Snout to pelvic fin origin | 47.6–52.2 | 30 | 49.5 | 46.9–51.5 | 20 | 49.5 |
| Snout to anal fin origin | 65.2–70.4 | 30 | 67.5 | 64.8–70.4 | 20 | 67.6 |
| Dorsal fin length | 23.5–27.6 | 30 | 25.0 | 20.9–26.6 | 20 | 24.5 |
| Anal fin base length | 21.6–27.8 | 30 | 24.9 | 22.1–26.6 | 20 | 24.4 |
| Orbit to dorsal fin origin | 39.5–45.3 | 30 | 42.4 | 40.0–44.7 | 20 | 42.7 |
| Head length | 24.3–26.9 | 30 | 25.3 | 23.6–26.5 | 20 | 25.0 |
| Percents of head length | ||||||
| Orbit diameter | 33.6–39.7 | 30 | 37.0 | 34.8–40.9 | 20 | 37.1 |
| Interorbital width | 29.3–33.7 | 30 | 31.5 | 29.9–36.6 | 20 | 32.4 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |
Bryconamericus misei
| Pedroso, Thiago Henrique, Deprá, Gabriel de Carvalho & Pavanelli, Carla Simone 2024 |
Bryconamericus aff. iheringii
| Cavalli D & Frota A & Lira AD & Gubiani EA & Margarido VP & Graca WJ 2018: 5 |