
Lebbeus longidactylus ( Kobjakova, 1936 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4175.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:54D34E46-687B-4FEB-AA66-F379A0FA9DB4 |
|
DOI |
https://doi.org/10.5281/zenodo.6080958 |
|
persistent identifier |
https://treatment.plazi.org/id/03BEA634-FFD9-FFA4-FF6F-F922FA5FFABD |
|
treatment provided by |
Plazi |
|
scientific name |
Lebbeus longidactylus ( Kobjakova, 1936 ) |
| status |
|
Lebbeus longidactylus ( Kobjakova, 1936)
[New Japanese name: Tsumenaga-ibara-mo-ebi] ( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Hetairus longidactyla Kobjakova, 1936: 191 (list), 194 (list), 222, figs 12, 13 (type locality: Sea of Okhotsk, 440–504 m); 1937: 110, fig. 13.
Lebbeus longidactyla .— Vinogradov 1950: 204 (key), 213, fig. 56.
Lebbeus longidactylus .— Hayashi 1992a: 108, 109 (list).—Komai et al. 2004: 122 ( Table 1), 123 (Table 2).— De Grave & Fransen 2011: 425.
Material examined. Nemuro Strait, off Rausu , Shiretoko Peninsula , eastern Hokkaido, 44°01.48’N 145°22.53’E to 44°02.31’N 145°23.55’E, 600–800 m, commercial shrimp trap, 14 July 2015, 1 ovigerous female (cl 23.5 mm), 1 male (cl 17.0 mm), CBM-ZC 13526 GoogleMaps ; same locality, 4 August 2014, 2 ovigerous females (cl 22.0, 18.6 mm), CBM-ZC 13527, 1 ovigerous female (cl 15.4 mm), AMF 1-04-01-01 GoogleMaps - 0024; same locality, 8 September 2014, 3 ovigerous females (cl 17.1, 17.7, 20.0 mm), AMF 1-04-01-01 -0025. GoogleMaps
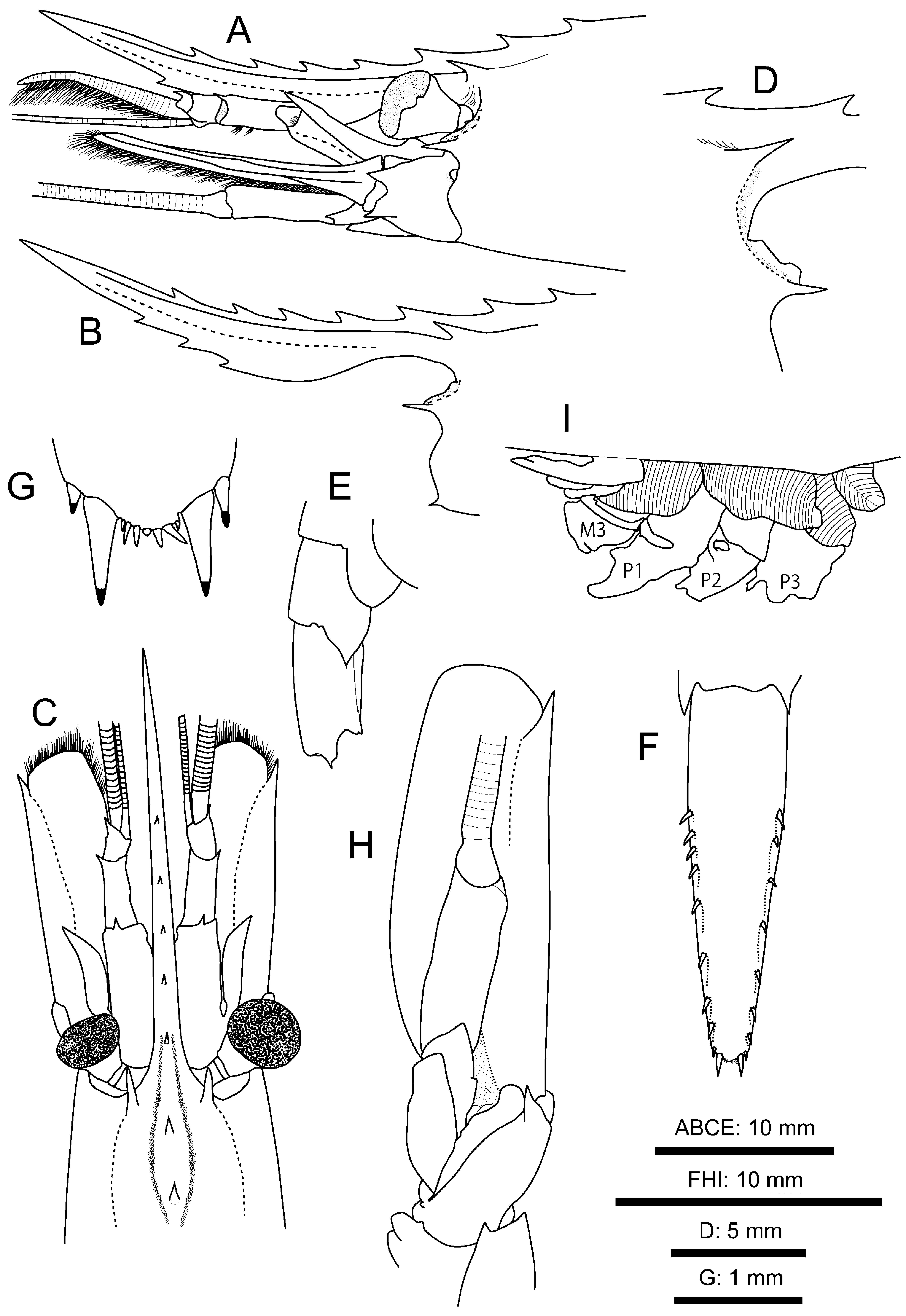
Redescription. Ovigerous females. Body ( Fig. 2 View FIGURE 2 A) relatively slender; integument thin, surface glabrous.
Rostrum ( Fig. 3 View FIGURE 3 A, B) gently curving dorsally, overreaching distal margin of antennal scale, 0.9–1.1 times as long as carapace; dorsal margin armed with 5–7 small spines including 3–5 on rostrum proper (anteriormost spine arising at about distal 0.7 of rostral length, and then anterior 0.3 leaving unarmed), and 2 postrostral (posteriormost spine located at about 0.2 of carapace length); ventral margin armed with 2–4 spines more widely spaced anteriorly; lateral carina obsolete.
Carapace ( Fig. 3 View FIGURE 3 A, B) with low postrostral ridge extending to 0.7–0.8 of carapace length, becoming broader and blunt posteriorly; orbital margin ( Fig. 3 View FIGURE 3 D) evenly concave, ventral part limbed marginally, postorbital region shallowly depressed just posterior to margin; supraorbital spine small, arising at level of posterior margin of orbit; no conspicuous notch below supraorbital spine; suborbital lobe distinct, rounded; antennal spine well developed, slender; pterygostomial spine small; anterolateral margin between antennal and pterygostomial spines gently sinuous.
Pleon ( Fig. 3 View FIGURE 3 E) with pleura of anterior four pleomeres rounded, unarmed, that of pleomere 5 with small posteroventral spine. Pleomere 3 rounded dorsally, posterodorsal margin somewhat produced posteriorly. Pleomere 6 about 1.6 times as long as pleomere 5, 2 times as long as high, with small posteroventral spine and sharply pointed posterolateral process. Telson ( Fig. 3 View FIGURE 3 F) moderately broad, armed with 7 or 8 pairs of dorsolateral spiniform setae; posterior margin terminating in small blunt triangular projection flanked by 2 pairs of unequal spiniform setae (mesial pair longer and stouter than lateral pair) ( Fig. 3 View FIGURE 3 G).
Eye ( Fig. 3 View FIGURE 3 A, C), subpyriform, cornea large, darkly pigmented, its maximum diameter about 0.2 of carapace length; ocellus absent. Eyestalk short, cup-shaped.
Antennular peduncle ( Fig. 3 View FIGURE 3 C, H) relatively slender. Article 1 longer than distal two articles combined, with slender spine on dorsolateral distal angle; stylocerite moderately slender, partially in touch with first article mesially, slightly curving laterally, terminating in acute spine reaching or slightly overreaching distal margin of article 1. Article 2 about 0.7 times as long as article 1, slightly broadened distally, armed with slender spine at dorsolateral distal angle. Article 3 short, with small spine on dorsodistal margin. Lateral flagellum ( Fig.3 View FIGURE 3 A) with thickened aesthetasc-bearing portion 0.5–0.6 times as long as carapace; mesial flagellum longer than lateral flagellum, 0.7–0.9 times as long as carapace.
Antenna ( Figs. 3 View FIGURE 3 A, C) with moderately stout basicerite, bearing blunt dorsolateral projection and sharp ventrolateral spine. Antennal scale 0.7–0.9 times as long as carapace, about 2.8 times as long as wide; lateral margin nearly straight; distolateral spine relatively slender, not reaching rounded distal margin of lamella. Carpocerite reaching midlength of antennal scale.
Mouthparts not dissected. Maxilliped 3 ( Fig. 4 View FIGURE 4 A) relatively slender, distinctly overreaching distal margin of antennal scale; ultimate article 3.1 times as long as penultimate article, bearing several darkly pigmented spiniform setae circumscribing terminal margin ( Fig. 4 View FIGURE 4 B); antepenultimate article slightly shorter than distal two articles combined, with 1 minute dorsolateral and 2 ventrolateral spiniform setae on distal margin ( Fig. 4 View FIGURE 4 C); exopod absent; coxa with strap-like epipod ( Figs. 3 View FIGURE 3 I, 4A).
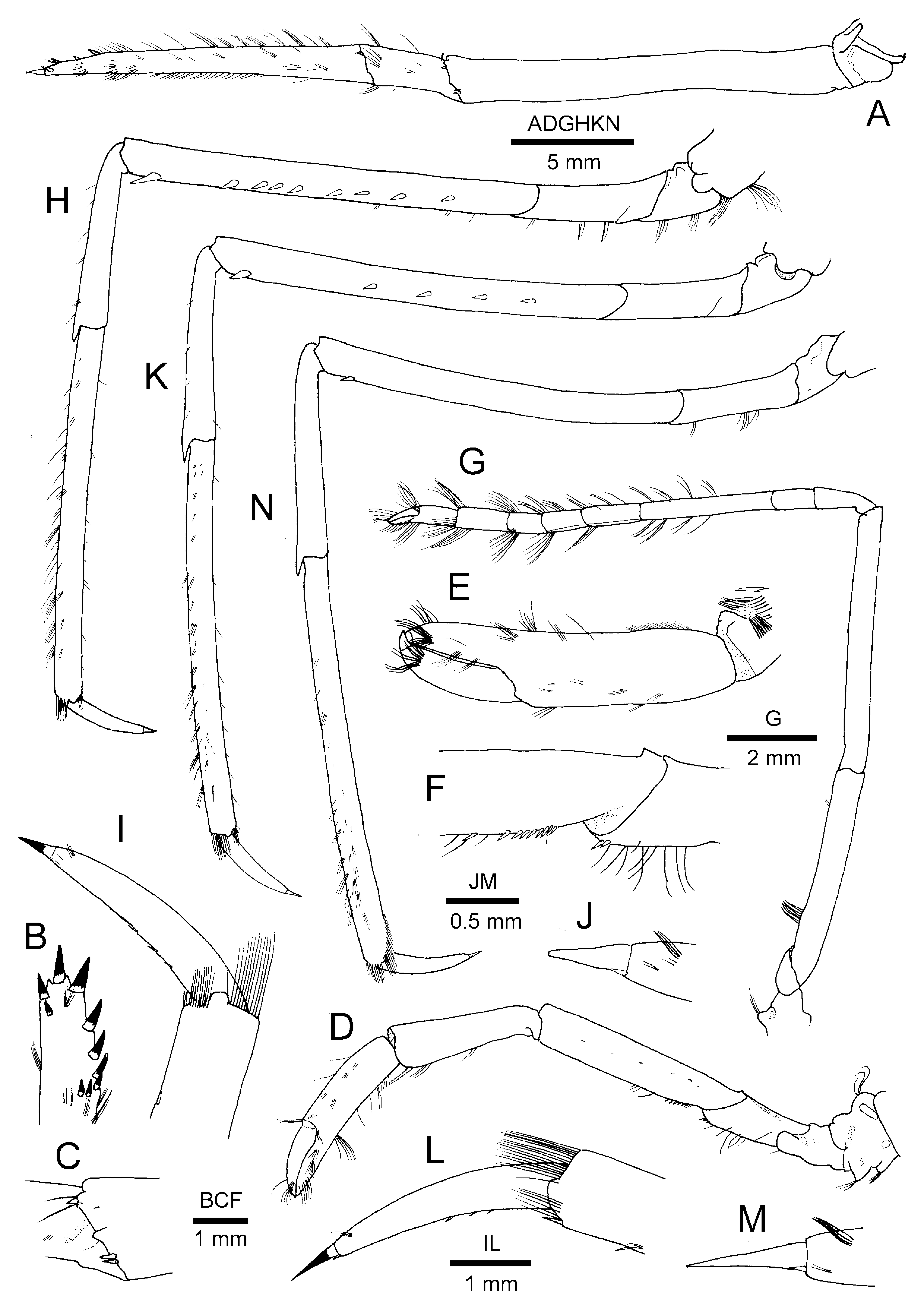
Pereopod 1 ( Fig. 4 View FIGURE 4 D) relatively slender. Chela ( Fig. 4 View FIGURE 4 E) 1.3 times as long as carpus; dactylus about 0.7 times as long as palm, terminating in 2 corneous claws; fixed finger terminating in single claw; palm about twice as long as wide. Carpus slightly widened distally, slightly longer than palm. Merus about 5.3 times as long as wide, with minute spine on dorsal margin proximally and short row of minute spiniform setae on ventral margin proximally. Ischium with few spiniform setae on ventrodistal margin.
Pereopod 2 ( Fig. 4 View FIGURE 4 G) moderately slender, overreaching antennal scale by length of chela. Chela about 0.2 times as long as carpus; dactylus 1.1–1.2 times as long as palm. Carpus subdivided into 7 articles, article 3 longest, occupying 0.3 times of whole carpal length; chela and distal 5 articles each with some tufts of setae. Merus about 1.2 times as long as ischium.
Pereopod 3 ( Figs. 4 View FIGURE 4 H) relatively slender, overreaching antennal scale by about half-length of propodus; dactylus ( Fig. 4 View FIGURE 4 I) slender, 0.3 times as long as propodus, about 6.9 times longer than wide, terminating in slender unguis ( Fig. 4 View FIGURE 4 J), bearing 3 minute, slender, subequal accessory spiniform setae on proximal 0.4–0.5 of flexor margin and 1 minute spiniform seta located at base of unguis; unguis clearly demarcated basally, 0.1–0.2 times as long as corps of dactylus, distal 0.7–0.8 darkly pigmented; propodus 13.0 times longer than wide; carpus 0.5 times as long as propodus, unarmed: merus 13.0 times longer than wide, armed with 5–9 spiniform setae on lateral surface.; ischium unarmed. Pereopod 4 ( Figs. 4 View FIGURE 4 K, L, M) similar to pereopod 3, overreaching antennal scale by 0.3 length of propodus; dactylus with 3 or 4 accessory spiniform setae; merus with 4–7 spiniform setae on lateral surface. Pereopod 5 ( Fig. 4 View FIGURE 4 N) generally similar to preceding pereopods, overreaching antennal scale by 0.2 length of propodus; dactylus with 3 or 4 accessory spiniform setae; propodus with cluster of grooming setae on flexor surface distally; merus with 1or 2 lateral spiniform setae.
Branchial formula summarized in Table 1. Epipods on pereopods 1 and 2 fairly reduced, each without terminal hook, corresponding setobranch on pereopod 2 and 3 absent; no epipods on pereopods 3–5. Coxa of pereopod 1 with setobranch corresponding to epipod on maxilliped 3.
Uropod with protopod terminating in acute spine posterolaterally; exopod subequal in length of endopod, either not reaching beyond posterior end of telson.
Eggs oval, large, 2.1–2.2 mm x 2.3–2.4 mm.
Male. Rostrum ( Fig. 5 View FIGURE 5 A) nearly straight, 1.1 times as long as carapace; dorsal margin with 7 spines including 2 postrostral, distal 0.4 unarmed; ventral margin with 3 spines. Antennular flagella more elongate and stouter than in females; lateral flagellum 1.2 times as long as carapace, thickened aesthetasc-bearing portion occupying 0.6 of whole length; inner flagellum ( Fig. 5 View FIGURE 5 A) 1.7 times as long as carapace. Pereopod 1 similar to that of females. Pereopods 2–5 similar to those of females; meri of pereopods 3–5 each with 6, 5, 1 spiniform setae on lateral surface. Endopod of pleopod 1 ( Fig. 5 View FIGURE 5 B) tapering into terminal appendix interna; mesial margin nearly straight, with row of curved minute spiniform setae; lateral margin sinuous. Appendix masculina of second pleopod ( Fig. 5 View FIGURE 5 C) shorter than appendix interna, non-tapered rod-like, bearing numerous long stiff setae on terminus to distal 0.7 of mesial face.
Maxillipeds Pereopods
1 2 3 1 2 3 4 5 Pleurobranchs – – – + + + + + Arthrobranchs – – – – – – – – Podobranchs – + – – – – – – Exopods + + – – – – – – Epipods + + + + + – – – Setobranchs – – – + – – – – Size. Male cl 17.0 mm, ovigerous females cl 15.4–23.5 mm.
Coloration in life. Body generally pink or light orange; pleomeres 1 to 3 each having obscure red transverse bands. Maxilliped 3 and pereopods generally red, with tinge of white on lateral surfaces of meri of pereopods 3–5. Pleopods 1–5 also reddish.
Distribution. Previously known only from the Sea of Okhotsk, at depths of 440– 504 m. The present material comes from the Nemuro Strait, southwestern part of the Sea of Okhotsk, at depths of 600–800 m, extending the bathymetrical range.
Remarks. Kobjakova (1936) diagnosed Lebbeus longidactylus in her key to species of the genus known from the Sea of Japan and the Sea of Okhotsk (as Hetairus ). No detailed information on the material she examined was provided, but it is apparent that the type material came from the Sea of Okhotsk at depths of 440–504 m from the summary of the geographical range of species treated in that paper (pp. 218–219). Subsequently, Kobjakova (1937) published a brief description of the species, indicating that two lots were available for study (the first lot from station 211, at depth of 443 m, containing two females, but the number of the specimens of the second lot from station 255, at depth of 504 m, was not specified). A holotype was not designated, and thus the original material should be considered syntypes . Vinogradov (1950) published a key to the species of Lebbeus from Far Eastern Russia. His account of L. longidactylus seems to be based on examination of the type material or type description by Kobjakova (1937), because the locality data on the species are identical. The present specimens agree well with those accounts in every diagnostic key character, particularly in the relatively long rostrum reaching or slightly overreaching the distal margin of the antennal scale, the possession of epipods on pereopods 1 and 2 (no epipod on pereopod 3) and the slender, distally simple, non-biunguiculate dactyli of pereopods 3–5. Although we have had no opportunity to access the type material, we identify our specimens with L. longidactylus with little hesitation because of the morphological consistency and the proximity of the collecting localities. The present material represents the first rediscovery of this rarely collected species since the original reports in 1936 and 1937.
Grouping of species within Lebbeus according to the number of pereopodal epipods was first applied by Rathbun (1904), and followed by many subsequent authors, e.g., Holthuis (1947), Butler (1980), Wicksten (1990), Hayashi (1992, 1993), Komai et al. (2004, 2012) and Chang et al. (2010). This grouping could be rather typological, but remains convenient for species identification and recognition. As mentioned above, L. longidactylus belongs to the group characterized by the possession of strap-like epipods on maxilliped 3 to pereopod 2, although the epipods on pereopod 1 and 2 is fairly reduced, lacking terminal hooks and corresponding setobranchs. Other than L. longidactylus , this species group includes the following 18 species ( Hayashi 1992a; Komai et al. 2004, 2012; Jensen 2006; Chang et al. 2010; McCallum & Poore 2010): L. acudactylus Jensen, 2006 (Alaska) , L. balssi Hayashi, 1992 (Japan and Korea), L. brandti ( Brashnikov, 1907) (northern Japan and Far Eastern Russia), L. brevirostris Chang, Komai & Chan, 2010 (Taiwan) , L. clarehannah McCallum & Poore, 2010 (Western Australia), L. compressus Holthuis, 1947 (Japan endemic, Pacific side), L. cristagalli McCallum & Poore, 2010 (Western Australia to Timor Sea), L. grandimana ( Brashnikov, 1907) (northern Japan and Far Eastern Russia), L. heterochaelus ( Kobjakova, 1936) (Sea of Okhotsk), L. kuboi Hayashi, 1992a (Sea of Japan), L.
laevirostris Crosnier, 1999 (Indonesia), L. laurentae Wicksten, 2010 (East Pacific Rise, hydrothermal vents), L. polaris ( Sabine, 1824) (Arctic and adjacent waters), L. scripssi Wicksten & Mèndez, 1982 (Gulf of California to Chile), L. splendidus Wicksten & Mèndez, 1982 (Peru) , L. unalaskensis ( Rathbun, 1902) (northern North Pacific), L. vinogradowi Zarenkov, 1960 (Sea of Okhotsk) and L. yaldwini Kensley, Tranter & Griffin, 1987 (New South Wales, Australia). As mentioned above, the dactyli of pereopods 3–5 in L. longidactylus are quite characteristic in the slender form with a simple terminus (non-biunguiculate) and poorly developed armature consisting of minute accessory spiniform setae restricted to the proximal 0.4–0.5 of the flexor margin. The vast majority of species of Lebbeus have terminally biunguiculate dactyli of pereopods 3–5, each having accessory spiniform setae distributed over the entire length of the flexor margin. Within the species group, only L. scripssi has similarly structured dactyli of pereopods 3–5. Nevertheless, L. longidactylus is readily distinguished from L. scripssi by the elongate rostrum (far overreaching the distal end of the antennular peduncle versus not reaching the distal margin of article 2 of the antennular peduncle) with more numerous dorsal spines (five to seven dorsal spines including two postrostral versus two to five including only one postrostral), the much more numerous dorsolateral spiniform setae on the telson (seven to eight versus three) and the presence of only one spine on the dorsodistal margin of article 1 of the antennular peduncle (vs. three spines) (cf. Wicksten & Méndez 1982). Lebbeus scrippsi is restricted to the eastern Pacific, ranging from the Gulf of California to Chile ( Wicksten & Méndez 1982; Hendrickx 2001).
Lebbeus unguiculatus Chang, Komai & Chan, 2010 (known from Sagami Bay, Japan to Taiwan) is also similar to L. longidactylus in the general structure of the dactyli of pereopods 3–5, although the former belongs to the species group having epipods on maxilliped 3 to pereopod 3. It further differs from L. longidactylus in the much shorter rostrum reaching only the distal margin of article 1 of the antennular peduncle, the presence of a deep notch just inferior to the base of the supraorbital spine (vs. such a notch being absent in L. longidactylus ), fewer dorsolateral spiniform setae on the telson (four or five pairs vs. seven or eight pairs), and the more numerous spination of the dorsodistal margin of article 1 of the antennular peduncle (single spine in L. longidactylus ).
Presently 67 species are known in Lebbeus , of which 31 species have been described only in the last two decades ( Fransen 1997; Komai 2002, 2011, 2013, 2015; Hanamura & Abe 2003; Komai & Takeda 2004; Komai et al. 2004, 2012; Jensen 2006; Ahyong 2009, 2010; Komai & Collins 2009; Komai & Komatsu 2009; McCallum & Poore 2010; Bruce 2010; Nye et al. 2012; Nye 2013; Schiaparelli et al. 2015; Matsuzaki et al. 2015), suggesting the existence of more unknown species still awaiting discovery. Thanks to the recent studies (e.g., Fransen 1997; Jensen 2006; Komai et al. 2012; Komai 2015) taxonomic ambiguity of some species has been substantially clarified, but as noted above, there are still several species represented only by insufficient type and/or subsequent descriptions, e.g., L. heterochaelus , L. profundus ( Rathbun, 1906) , L. spinirostris , L. uschakovi , L. vicinus ( Rathbun, 1902) , L. vinogradowi , and L. washingtonianus ( Rathbun, 1902) . Further study using type material and efforts to collect additional samples will be necessary to clarify the taxonomic identities of these enigmatic taxa.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lebbeus longidactylus ( Kobjakova, 1936 )
| Komai, Tomoyuki, Matsuzaki, Koji & Hibino, Mai 2016 |
Lebbeus longidactylus
| De 2011: 425 |
| Hayashi 1992: 108 |
Lebbeus longidactyla
| Vinogradov 1950: 204 |
Hetairus longidactyla
| Kobjakova 1936: 191 |