
Ziminella vrijenhoeki, Valdés & Lundsten & Wilson, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4526.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:3CFFF3AC-C447-4FCE-B6F8-D2B7BAE8B678 |
|
DOI |
https://doi.org/10.5281/zenodo.5971404 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87CC-FFA8-DE1D-9B8E-9D03790E5B1B |
|
treatment provided by |
Plazi |
|
scientific name |
Ziminella vrijenhoeki |
| status |
sp. nov. |
Ziminella vrijenhoeki View in CoL sp. nov.
( Figs. 2 View FIGURE 2 D–E, 11–13)
Type material. Holotype: Blue whale skeleton (implanted on 5 Oct 2004), Monterey Bay, California (36.772, - 122.083), 1018 m depth, ROV Tiburon (dive 1072), 13 Jan 2007, 20 mm preserved length ( SIO-BIC M12137 View Materials ), GenBank accession numbers: MH 756136 View Materials ( COI), MH 756130 View Materials (16S), MH 756141 View Materials (H3).
Paratype: Whale skeleton (unknown species, likely naturally occurring), Monterey Bay, California, (36.7721, - 122.0831), 595 m depth, ROV Doc Ricketts (dive 94), 16 Nov 2009, 24 mm preserved length, dissected ( SIO-BIC M12135 View Materials ), GenBank accession numbers: MH 756135 View Materials ( COI), MH 756129 View Materials (16S) , MH 756140 View Materials (H3).
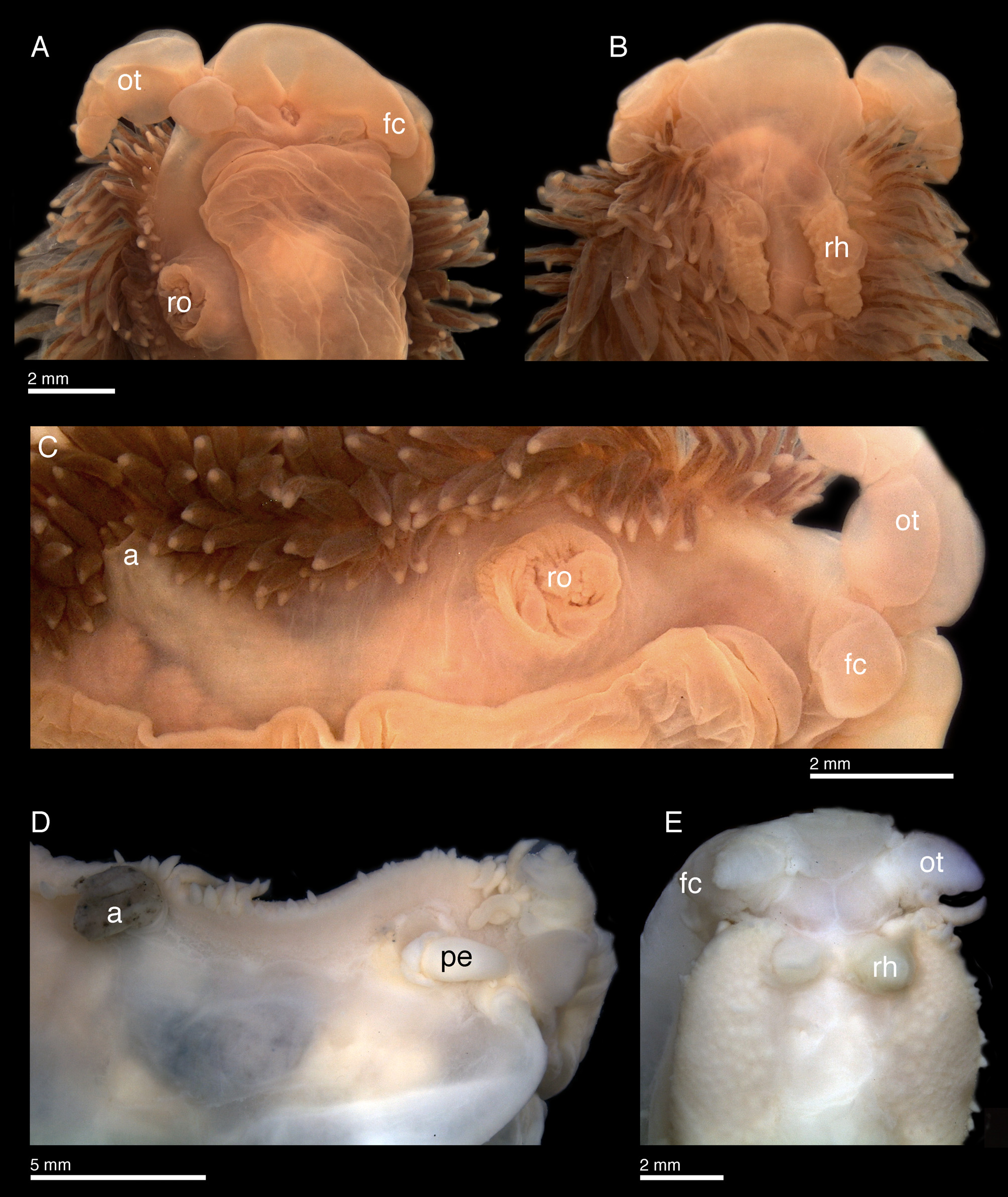
Description. Body elongate, narrow ( Figs. 2D View FIGURE 2 ), with numerous elongate, thin, dorso-lateral cerata. Cerata arranged in densely packed rows, running from behind oral tentacles to posterior end of body ( Figs. 2D View FIGURE 2 , 11 View FIGURE 11 C–D). Cerata increase in size in each row, lateral cerata typically shorter, dorsal cerata longer. Oral tentacles short and wide ( Figs. 11 View FIGURE 11 A–D, 11E), nearly conical in live animal ( Fig. 2D View FIGURE 2 ). Foot corners tentacular, short, wide, almost spherical ( Figs. 11C, 11D View FIGURE 11 ). Rhinophores elongate, smooth ( Figs. 2D View FIGURE 2 , 11B, 11E View FIGURE 11 ). Reproductive opening on right anterior portion of the body ( Fig. 11 View FIGURE 11 C–D), penis smooth and elongate ( Fig. 11D View FIGURE 11 ). Anal opening dorso-lateral, posterior to pericardium ( Fig. 11 View FIGURE 11 C–D). Body color cream to reddish-orange, semi translucent, with viscera visible through skin as a yellowish mass ( Fig. 2 View FIGURE 2 D–E). Oral tentacles the same color as body, rhinophores redder and darker. Cerata translucent pale orange, with red digestive branches and white cnidosacs.
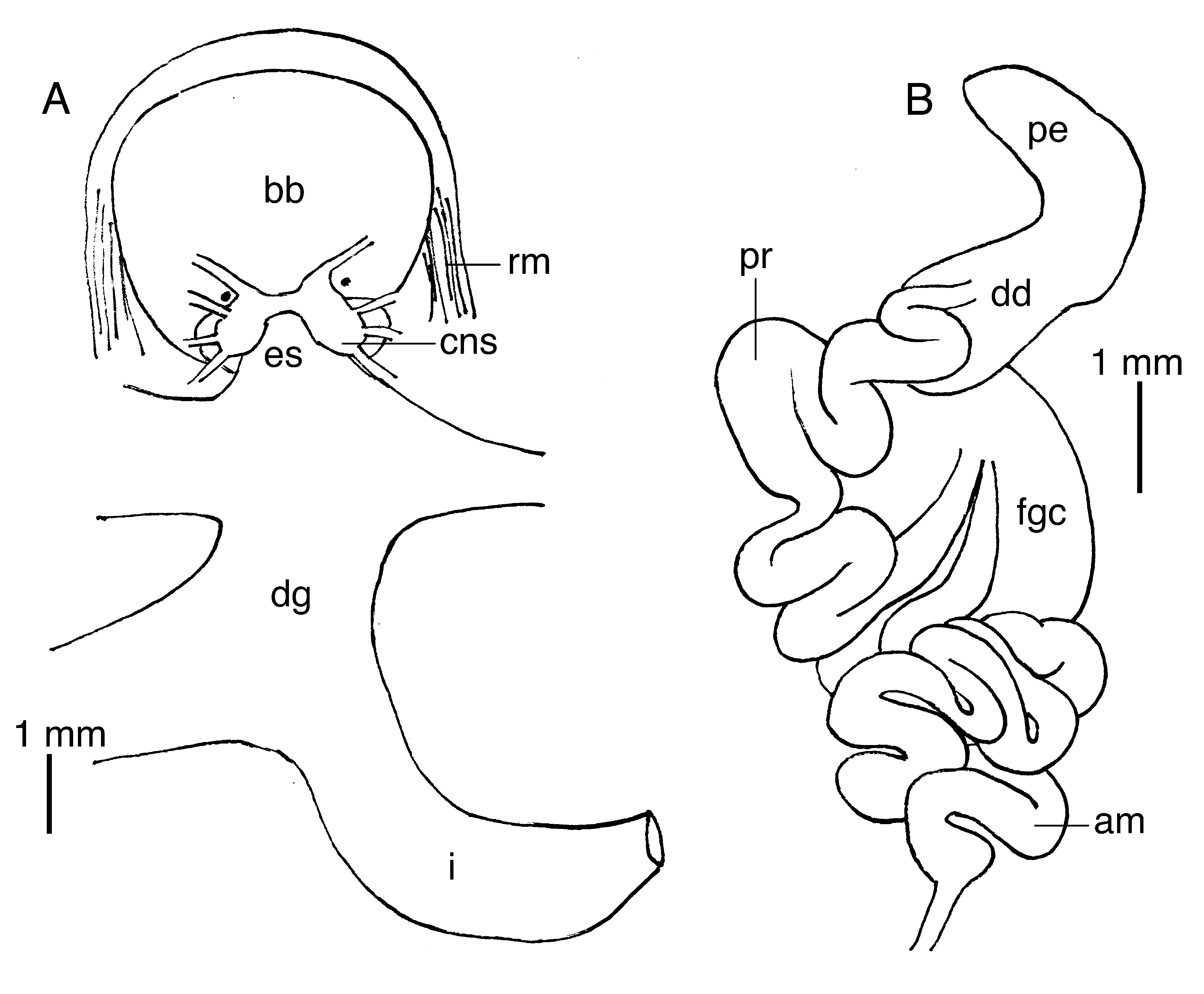
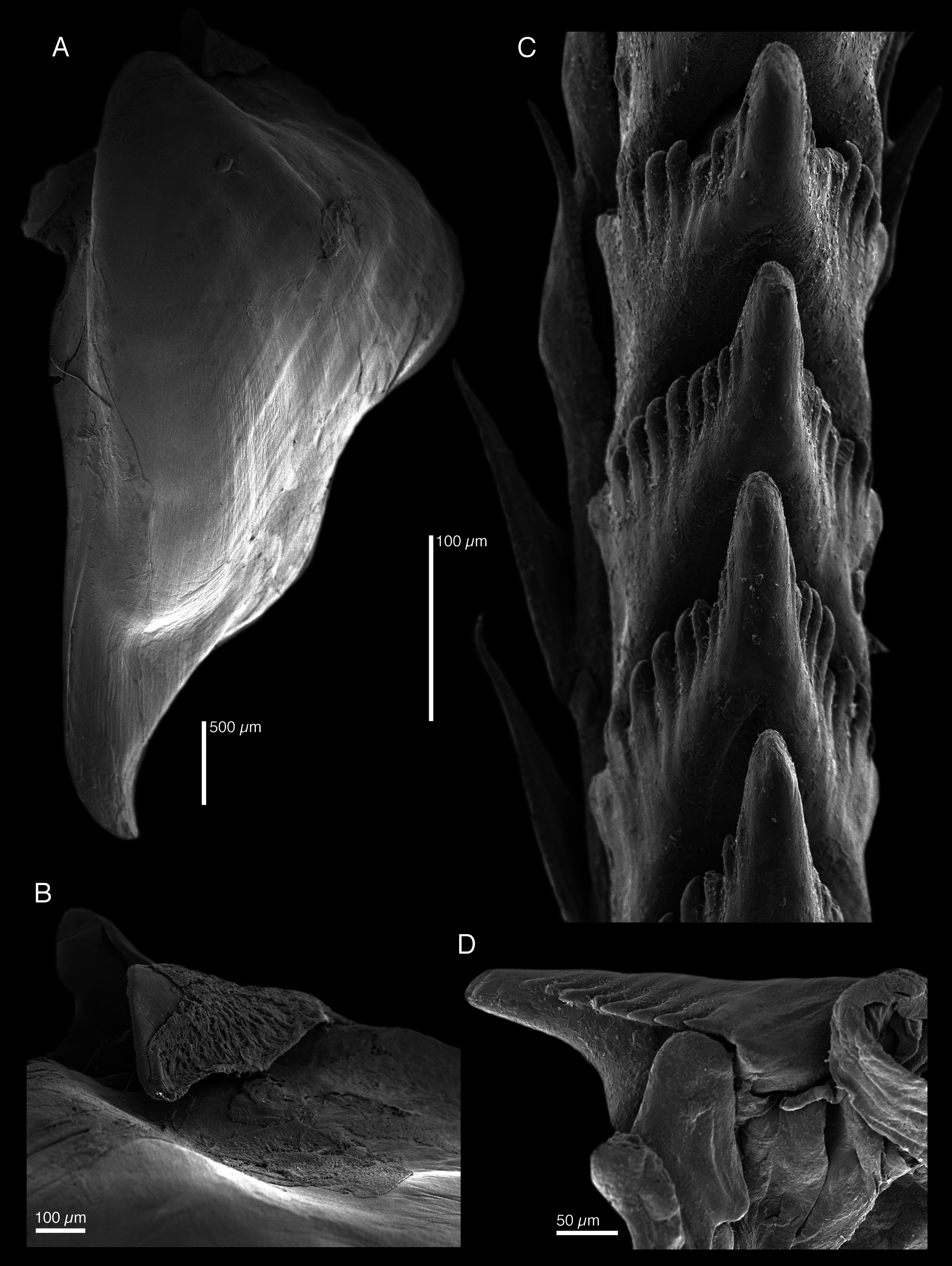
Digestive system with large, muscular buccal bulb ( Fig. 12A View FIGURE 12 ). Esophagus narrow, short, connecting anteriorly into buccal bulb. Digestive gland with lateral branches entering cerata. Intestine emerging laterally from right side of digestive gland, forming a loop and opening into anus on right side of body. Radular formula 27 × 1.1. 1 in paratype (SIO-BIC M12135 View Materials ). Rachidian radular teeth narrow ( Fig. 13C View FIGURE 13 ), with arch-shaped base and a single central, blunt cusp. Each tooth with 6–8 blunt denticles on each side of cusp ( Fig. 13D View FIGURE 13 ). Lateral teeth triangular, elongate, with numerous minute denticles on inner side. Jaws elongate ( Fig. 13A View FIGURE 13 ), no denticles on masticatory border (13B).
Reproductive system with very elongate, convoluted ampulla forming numerous loops ( Fig. 12B View FIGURE 12 ), opening into female gland complex next to prostate opening. Prostate tubular, very elongate, forming numerous loops, opening into elongate, muscular deferent duct. Penis simple, elongate. No vagina nor bursa copulatrix were observed, probably an artifact of the dissection.
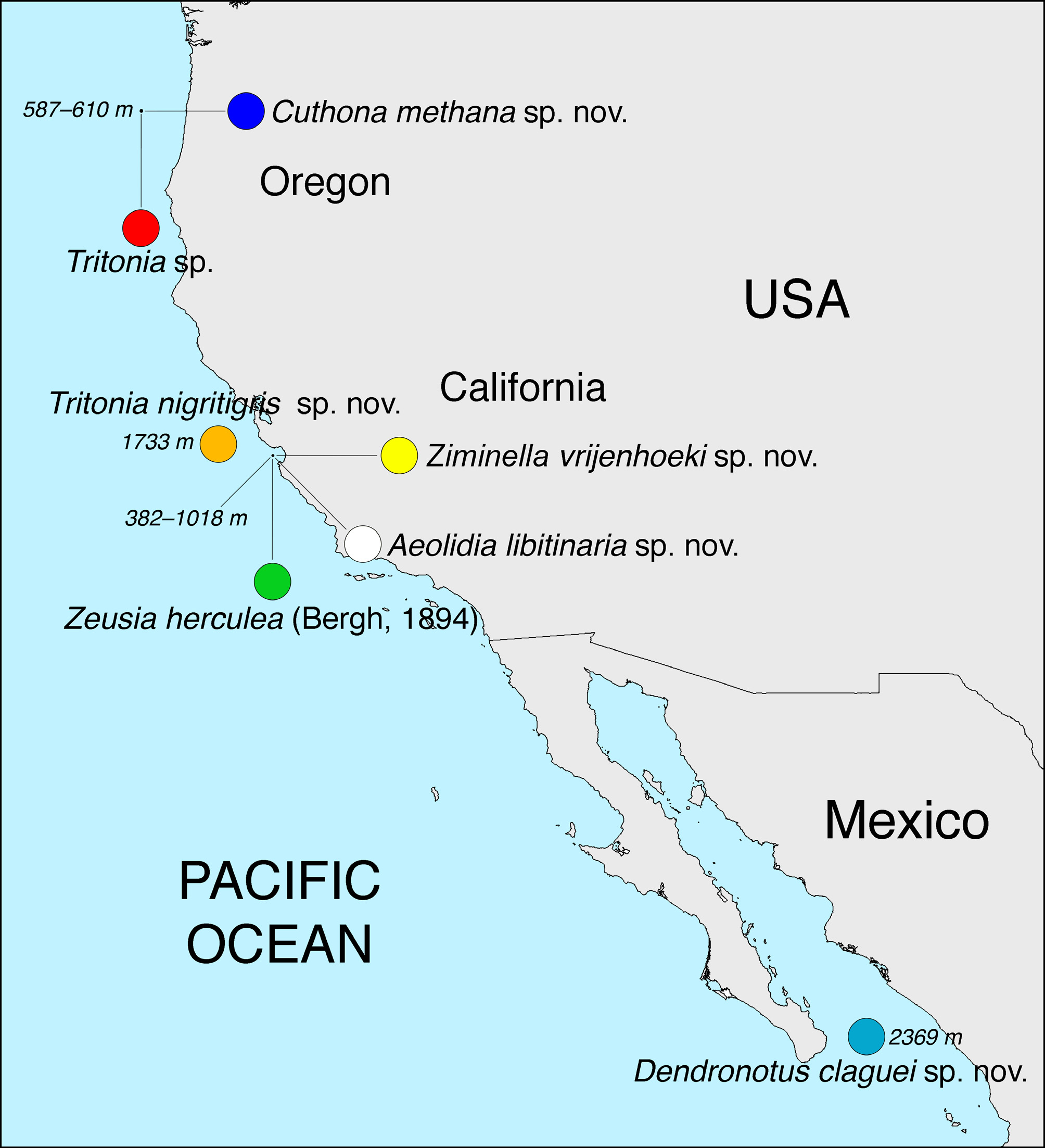
Biology. This species was collected at two different whale-falls in Monterey Canyon at depths of 1018 m (holotype) and 595 m (paratype) in Monterey Bay, California ( Fig. 1 View FIGURE 1 ). The holotype was collected from a vertebra of an implanted Blue Whale carcass that was being surveyed as part of a time-series analysis of whale-fall ecology ( Lundsten et al. 2010b). The whale skeleton where the holotype was collected was in the sulphophilic stage of degradation with dense colonies of Osedax sp. (siboglinid polychaete), additional unidentified polychate worms, Tritonia tetraquetra (nudibranch), Neptunea -Buccinum complex (gastropods), Chionoecetes tanneri (crabs), the fishes Sebastolobus sp. and Lycodapus sp. and Vesicomyidae (chemosynthetic bivalves). The paratype was collected from the sediment surface in the vicinity of a presumed natural whale carcass perched upon the edge of the Monterey Canyon, with some bones of the carcass spread down a steep cliff. Two specimens were observed and recorded onto video tape during this dive (Doc Ricketts dive 94), however, only one specimen was collected.
Both specimens were observed upon the sediment. A subtle color variation was noticed between the D94 observations, with the first being a darker, bluer color ( Fig. 2E View FIGURE 2 ) and the second, which was collected, being more of a reddish orange ( Fig. 2D View FIGURE 2 ). This whale was presumed to be in the early-stages of decomposition with few sulphophilic organisms or Osedax sp. (siboglinid polychaete) worms observed, although flesh was completely removed. Other organisms observed nearby included the fishes Merluccius productus and Sebastolobus sp., the echinoderms Asteronyx sp., Rathbunaster californicus , Pterasteridae , Poraniopsis inflata and the polychaetes Sabellidae and Serpulidae , as well as Eualus macrophthalmus (caridean shrimp), and Caprellidae (amphipods).
Phylogenetic position. Based on the phylogenetic analyses here conducted ( Fig. 14 View FIGURE 14 ), Ziminella vrijenhoeki sp. nov. is nested in a polytomy containing Ziminella abyssa Korshunova et al., 2017 and a clade including Ziminella salmonacea ( Couthouy, 1838) and Ziminella circapolaris Korshunova et al., 2017 .
Etymology. The species is named in honor of Robert C. Vrijenhoek, MBARI Senior/Adjunct Evolutionary Biologist. Bob was the Chief Scientist of MBARI’s whale-fall time series surveys, during which these specimens were collected.
Remarks. Korshunova et al. (2017) placed Flabellina salmonacea ( Couthouy, 1838) and the new species Ziminella abyssa Korshunova et al., 2017 and Ziminella circapolaris Korshunova et al., 2017 in the new genus Ziminella Korshunova et al., 2017 . Ziminella constitutes a well-supported clade, and in the analyses conducted by Korshunova et al. (2017) it is nested in a larger, equally well-supported clade. This larger clade, which contains other species previously assigned to Flabellinidae , including the type species of Flabellina , F. affinis (Gmelin, 1791) , was not recovered in the analyses here conducted. The decision of Korshunova et al. (2017) to split Flabellina into a number of new genera dramatically increases the taxonomic complexity of the group, makes the placement of species not studied genetically more difficult, and created taxa supported by few synapomorphies. Additionally, several of those clades were not recovered in our analyses, indicating that further work is needed to produce a reliable phylogeny and classification for aeolid nudibranchs. However, based on the results of the analyses here conducted, which clearly show the close relationship between Ziminella vrijenhoeki sp. nov. and other species assigned to Ziminella by Korshunova et al. (2017), we provisionally place this new species in the genus Ziminella , until more reliable phylogenies become available.
Based on Korshunova et al. (2017) and the results of the analyses presented here, the closest species to Ziminella vrijenhoeki sp. nov. are Z. salmonacea , originally described from Charles River, Massachusetts ( Couthouy, 1838) and subsequently reported from different localities in the North Atlantic ( Kuzirian 1979), Z. abyssa , originally described from deep water (2676 m depth) in the Sea of Japan (Korshunova et al., 2017), and Z. circapolaris , originally described from Franz Josef Land, Arctic Ocean (Korshunova et al., 2017). Ziminella salmonacea has also been reported from the North Pacific ( Wakeling 2002), but Martynov (2013) proposed the Pacific Ocean animals belong to Flabellina japonica (Volodchenko 1941) .
According to Korshunova et al. (2017) species of Ziminella are distinguished by their penial morphology, Z. salmonacea possesses a slightly folded penis, whereas Z. abyssa and Z. japonica have conical penises; but no data is provided for Z. circapolaris . The penis of Z. vrijenhoeki is folded and therefore more similar to that of Z. salmonacea . Ziminella vrijenhoeki sp. nov. and Z. salmonacea are genetically and morphologically distinct, the radula of Z. salmonacea as described by Kuzirian (1979) has a much more elongate rachidian tooth, with smaller denticles. Although we had no access to sequence data of F. japonica for comparison, this species is clearly different from Z. vrijenhoeki sp. nov. The body of F. japonica is cream to pinkish externally and the digestive branches in the cerata are cream ( Behrens & Hermosillo 2005) whereas Z. vrijenhoeki sp. nov. is reddish-orange, and the cerata are translucent pale orange, with red digestive branches and white cnidosacs.
The single specimen of Ziminella circapolaris Korshunova et al., 2017 included in the analyses here presented is nested within Z. salmonacea raising doubts on the validity of the former species.


| MH |
Naturhistorisches Museum, Basel |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heterobranchia |
|
Order |
|
|
SubOrder |
Cladobranchia |
|
Family |
|
|
Genus |