
Odontholariopsis Audisio & Cline, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.5319334 |
|
DOI |
https://doi.org/10.5281/zenodo.5342840 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87CC-F667-FF94-BA5E-FE51FB9FFD77 |
|
treatment provided by |
Felipe |
|
scientific name |
Odontholariopsis Audisio & Cline |
| status |
gen. nov. |
3. Odontholariopsis Audisio & Cline , gen. nov.
( Figs. 3 a–h View Fig )
Type species. Meligethes nebulosus Reitter, 1872: 247 (by present designation) [= Odontholariopsis nebulosus (Reitter, 1872) comb. nov.].
Generic description and diagnosis. Inclusive species vary greatly in size (1.4–2.8 mm length), and share the following combination of characters.
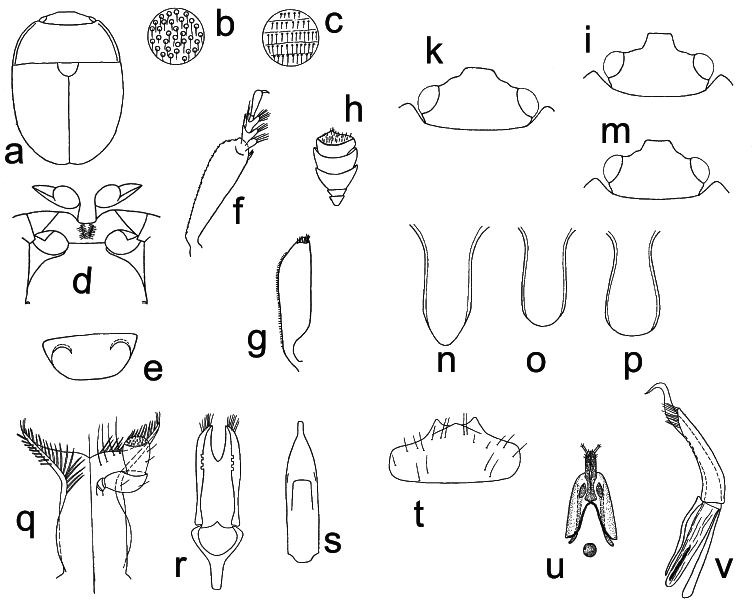
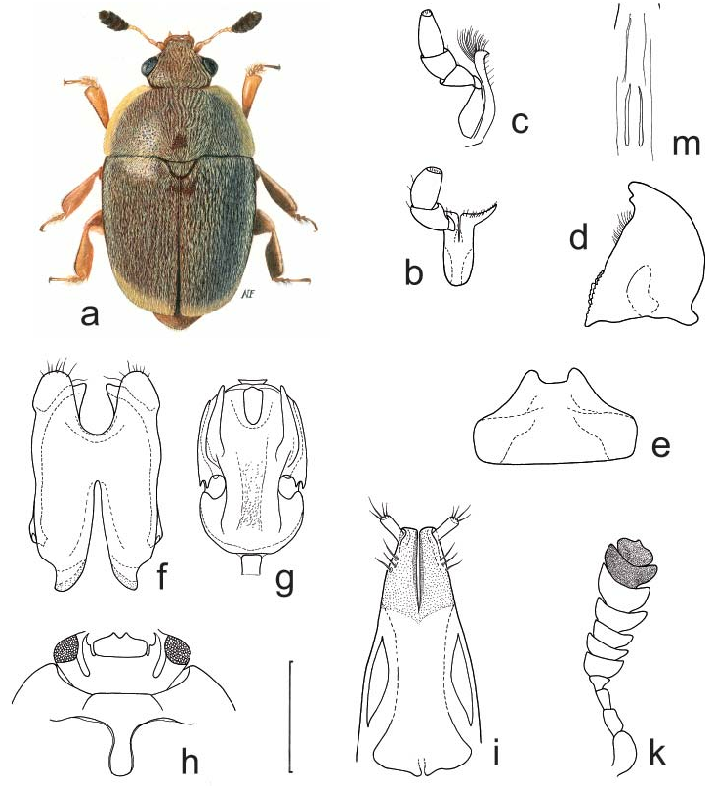
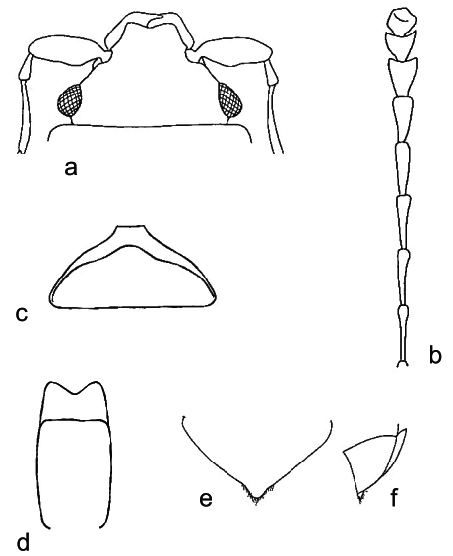
Body color and pubescence: pubescence golden to silvery-whitish or brownish, short and fine, recumbent, sparse, never obscuring the variably colored (reddish, brown, blackish, or brown with yellowish elytra: Figs. 3a View Fig ; Figs. 129–130 in KIREJTSHUK & AUDISIO (1995)) dorsal body surface; pronotal and elytral sides narrowly flattened, typically the same color as disk. Lateral margin of pronutum and elytra with a series of usually distinct setae, each seta 0.7–0.8× as long as those on elytral disc; posterior margin of pronotum comprising moderately long, distally trifid or tetrafid microsetae, microsetae uniformly also along middle region anterior to scutellum ( Fig. 3e View Fig ).
Dorsal habitus: body moderately convex, variably shaped, usually moderately short and wide, oval, in some species exceptionally short and wide ( Figs. 3a View Fig ; Figs. 6–9 View Fig View Fig View Fig View Fig and 129–130 in KIREJTSHUK & AUDISIO 1995); dorsal punctures on discal portion of pronotum as large as or larger than eye facets, shallowly impressed and densely distributed; anterior margin of clypeus slightly arcuately emarginate, simple, i.e. always without small distinct medial bulge, and not distinctly bordered ( Fig. 3b View Fig ), with circum-ocular furrows (occipital sulci) on dorsal side of head absent ( Fig. 3b View Fig ); eyes large and usually moderately projecting laterally ( Figs. 3a, b View Fig ); pronotum with faintly distinct posterior angles, rounded to obtuse and never directed posteriorly ( Fig. 3a View Fig ); scutellum regularly punctured in most of exposed portion; elytra with simple punctation, never transversely strigose; elytral humeral angle moderately distinct, not protruding laterally ( Fig. 3a View Fig ); elytral humeral striae usually not distinct; elytral pre-sutural striae visible, originating at scutellar vertex, terminating at elytral apex, and delimiting a faintly distinct, flat, not raised sutural border on each elytron, border widest at posterior third, slightly narrower than proximal width of 3 rd antennomere; elytral apices truncately rounded in both sexes ( Fig. 3a View Fig ); pygidium partially exposed, moderately convex, apically rounded in both sexes ( Fig. 3a View Fig ; Figs. 6–9 View Fig View Fig View Fig View Fig and 129–130 in KIREJTSHUK & AUDISIO 1995).
Ventral habitus: antennal furrows markedly delimited, nearly parallel-sided, or slightly divergent posteriorly; mentum subpentagonal ( Fig. 3c View Fig ); prosternal antennal furrows of anterior margin of prosternum scarcely raised and short, but distinct ( Fig. 3c View Fig ); prosternal process relatively narrow, but with strongly dilated subapical portion 2.3–2.5× as wide as maximum width of 1 st antennomere, apex blunt ( Fig. 3f View Fig ); lateral borders of prosternal process delimiting shallowly impressed but distinct furrows, distally terminating before predistal lateral expansions ( Fig. 3f View Fig ); posterior margin of mesoventrite simple, never medially incised ( Fig. 3f View Fig ); moderately expressed sexual dimorphism on metaventrite, with variably developed impressions; first two visible abdominal ventrites simple in both sexes, without tufts of setae; caudal marginal lines of metacoxal cavities simple, parallel and contiguous to posterior margin of metacoxal cavities, without arched impression of outer ‘axillary’ line (as in Fig. 4k View Fig ); ‘axillary’ space on first abdominal ventrite moderately developed, ‘axillary’ angle widely obtuse (as in Fig. 4k View Fig ); small, short, and shallowly impressed arched impressions on basal portion of last visible abdominal ventrite mostly concealed by distal portion of penultimate visible abdominal ventrite ( Fig. 3h View Fig ).
Appendages: male 1 st antennomere 0.8–1.0× as long as protibial width excluding distal teeth ( Figs. 3a, c View Fig ); 3 rd antennomere in both sexes peculiarly short, usually only 1.7–1.8× as long as wide, and nearly 0.7× as long as but distinctly thinner than 2 nd antennomere ( Figs. 3a, c View Fig ); 4 th and 5 th antennomeres in both sexes subequal, short, nearly as long as wide; antennal club compact, small, simple, comprising last 3 antennomeres in both sexes (8 th antennomere moderately widened, nearly 0.7× as wide as 9 th antennomere) ( Figs. 3a, c View Fig ), much narrower than width of protibiae, not sexually dimorphic; labial palpi short in both sexes ( Fig. 3c View Fig ), terminal segment 1.4–1.5× as long as wide; maxillary palpi moderately long and slender in both sexes ( Fig. 3c View Fig ), terminal segment 1.9–2.0× as long as wide; mandible small-sized ( Fig. 3c View Fig ), apex typically acuminate, not sexually dimorphic; tarsal claws simple, never toothed at base ( Fig. 3d View Fig ); tarsi of normal size and shape, 0.6–0.7× as long as corresponding tibiae ( Figs. 3a, d View Fig ); protibiae with a series of usually moderately large, uneven, scarcely acuminate teeth on lateral margin ( Figs. 3a View Fig ; Figs. 78–80 in KIREJTSHUK & AUDISIO 1995); meso- and metatibiae on lateral margin bearing an apparently simple and usually even row of large and robust spurs, with peculiar U-shaped sinuosity at distal third, usually armed by additional outer spur ( Fig. 3d View Fig ; Figs. 91–92 in KIREJTSHUK & AUDISIO 1995); meso- and metatibiae of variable width, usually moderately wide and flat ( Fig. 3a View Fig ; Figs. 6–9 View Fig View Fig View Fig View Fig and 91–92 in KIREJTSHUK & AUDISIO 1995), never distinctly subtrapezoidal or axe-shaped; tarsal plates of prolegs moderately wider in males; strongly manifested sexual dimorphism in meso- and metafemoral shape, metafemoral margins in males bearing a single large, triangular or finger-like projection, reduced to a faintly distinct gibbosity in females ( Fig. 3g View Fig ; Figs. 100, 102– 104 in KIREJTSHUK & AUDISIO 1995).
Male genitalia: processes along inner side of parameres absent ( Figs. 25–26 View Fig View Fig and 39 View Fig –44 in KIREJTSHUK & AUDISIO 1995), usually with shallow excision along distal margin, without deep median longitudinal desclerotization from proximal portion of tegmen and extending to medial distal V-shaped excision; median lobe of aedeagus variable, without lateral emargination, rounded, subtruncate to obtusely emarginate distally.
Female genitalia (ovipositor): variably shaped, usually small; styli usually long and relatively large, simple, cylindrical, moderately pigmented, inserted close to apex of contiguous gonostyloids; each gonostyloid lightly sclerotized and more darkly pigmented distally, with a simple, never indentate outer portion of basicoxites (Figs. 48–51 in KIREJTSHUK & AUDISIO 1995), and a single, variably shaped, pigmented and moderately sclerotized arcuate area along outer subdistal portion of gonostyloids. ‘Central point’ of ovipositor usually centrally located, without proximad directed spicule.
Etymology. The generic name is derived from a combination of the Greek ‘οδούς, οδοντος’ (= tooth), and Lariopsis , to emphasize the peculiarly shaped and toothed posterior margin of meso- and metafemora in males, and its placement within the Lariopsis generic complex. Gender masculine.
Biology. All species strictly associated for larval development with inflorescences of Mesembryanthemaceae , especially Carpobrotus N. E. Br. , Cephalophyllum (Haw.) N. E. Br. , Ruschia Schwant. , Lampranthus N. E. Br. , Delosperma N. E. Br. , and allied genera ( KIREJTSHUK & AUDISIO 1995, and unpublished data).
Phylogenetic position. Odontholariopsis gen. nov. is likely more closely related to Asterogethes gen. nov., and less so to Lariopsis and Neolariopsis gen. nov., but forming with them the newly defined Lariopsis generic complex, which is supported with evidence from both adult morphology and molecular datasets ( TRIZZINO et al. 2009). Phylogenetic relationships of Odontholariopsis gen. nov. with Acanthogethes , Clypeogethes , and other ancestral Meligethinae genera remain unclear, and are only weakly supported by molecular data.
Taxonomy and geographic distribution. Odontholariopsis gen. nov. includes four described species distributed in Southern Africa ( KIREJTSHUK & AUDISIO 1995).
Odontholariopsis aurimaculatus ( Kirejtshuk & Audisio, 1995) comb. nov. South Africa: W Cape Odontholariopsis haagii (Reitter, 1872) comb. nov. South Africa: W Cape, S Namibia Odontholariopsis nebulosus (Reitter, 1872) comb. nov. South Africa: W Cape Odontholariopsis sphaeroideus ( Kirejtshuk & Easton, 1988) comb. nov. South Africa: W and E Cape






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.