
Cardiodactylus tankara, Robillard, 2009
|
publication ID |
https://doi.org/10.5252/z2009n3a11 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDFA31-9D16-FF87-3E3D-FB2FFB47B01C |
|
treatment provided by |
Marcus |
|
scientific name |
Cardiodactylus tankara |
| status |
sp. nov. |
Cardiodactylus tankara View in CoL n. sp.
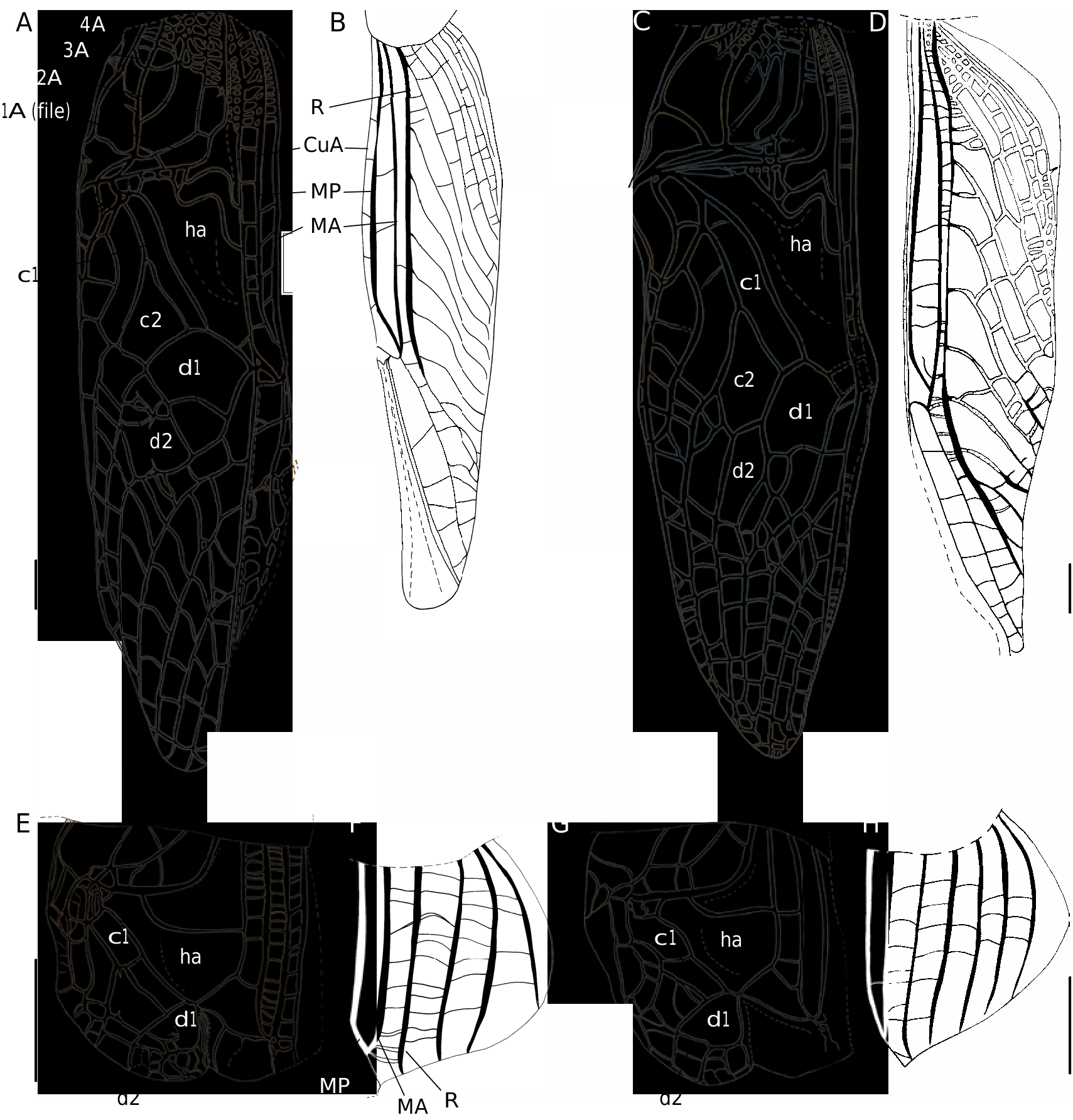
( Figs 1C, D View FIG ; 3A View FIG ; 4 View FIG F-H; 5B; 6B; 7J-L; 8; 15; 17D, E)
Cardiodactylus sp. – Robillard et al. 2007: 1254 (calling song analysis).
TYPE MATERIAL. — Holotype: Vanuatu. [Province Sanma], Espiritu Santo Is., Butmas , forêt du plateau de Tankara, 15°21’56’’S, 166°59’E, 600 m alt., 13.X.2006, jour, 12AM, ♂ (TR-1), sur plante de sous-bois, sur tronc à h = 2 m, forte pente, enregistrement (chant d’appel) TR-take 16, photo + vidéo site SH, T. Robillard ( MNHN- ENSIF2409 ). GoogleMaps
Allotype: same locality and collector as holotype, 16.X.2006, nuit, 6-10PM, ♀ (TR-128), sur plante de sous-bois, vidéo TR-V4 de ponte en semi-captivité, obs. copulation avec ♂ (TR-127) (MNHN-ENSIF2410).
Paratypes ( 9 ♂♂; 2 ♀♀): same locality and collector as holotype, X.2006, adulte en élevage, souche F0, élevage Santo TR, 1 ♀ (MNHN-ENSIF2432) ; 13.X.2006, jour, 7-11AM, 1 ♂ (TR-57), sur plante de sous-bois, sur fougère à h = 1 m, enregistrement en semi-captivité (chant d’appel) TR-takes 24, 26-30; 13.X.2006, jour, 12AM, 1 ♂ (TR-2), sur plante de sous-bois, sur fougère, fauchage, h = 1,5 m, enregistrement TR-takes 17-19; 14.X.2006, jour, 12AM, 1 ♂ (TR-70), sur plante de sous-bois, fougère h = 2,5 m, photo SH, obs. chant d’appel; 16.X.2006, nuit, 6-10PM, 1 ♂ (TR-122), sur plante de sous-bois; 16.X.2006, nuit, 1 ♂ (TR-125), sur plante, vidéo TR-V1, 1 ♂ (TR-126), sur plante, 1 ♂ (TR-145), sur plante de sous-bois h = 2 m, 1 ♂ (TR-130), sur plante de sous-bois/fougère h = 1,5 m, terrain en pente et 2 ind. proches, enregistrement en milieu naturel (chant d’appel), TR-take 22, 1 ♂ (TR-128’, alcool), sur plante, PIII: molec TR; 17.X.2006, nuit, 1 ♀ (TR-145’), sur plante, souche F0 élevage TR Santo , 1 ♂ (TR-146), herbes hautes en bord de piste, sur fougère h = 1,5 m, chant d’appel: vidéo SH (MNHN-ENSIF2411-2414, 2416-2422) .
TYPE LOCALITY. — Vanuatu, Espiritu Santo Is., plateau of Tankara, near the village of Butmas.
OTHER MATERIAL EXAMINED. — Vanuatu. [Province Sanma], Espiritu Santo Is., Butmas, forêt du plateau de Tankara , 15°21’56’’S, 166°59’E, 600 m alt., 16.X.2006, nuit, 6-10PM, 2 ♀♀ juv. (TR-123, 131), sur plante de sous-bois GoogleMaps ; 1 ♂ (TR-124), sur plante de sous-bois h = 1,8 m, vidéo TR-V2 5h30PM avant la tombée de la nuit; 17.X.2006, nuit, 1 juv. (TR-142), 1 juv., forêt de brume, sur plante de sous-bois h = 0,8 m feuille de liane, T. Robillard (MNHN-ENSIF2415, 2423-2425, 2566). — Butmas, Grotte de Fapon /sentier, 15°19’51,6’’S, 166°57’53,6’’E, forêt dans la doline, 15.X.2006, jour, 1 juv. (TR-78), sur plante GoogleMaps ; 15.X.2006, jour, 10-12AM, 1 juv. (TR-114), zone ouverte, sur plante de sous-bois h = 0,5 m, 1 ♀ juv. (TR-77), sur plante, PIII molec.TR, T. Robillard (MNHN-ENSIF2426-2428) — 13.X.2006, nuit, 1 ♂ juv. (nº 22), sur plante de sous-bois, L. Desutter- Grandcolas (MNHN-ENSIF2429).— Reared specimens: F1, 2006-2007, 4 ♂♂ (2 recorded, MNHN-ENSIF2430, 2431), 1 ♀ ; F2, 2007-2008, 1 ♂ (recorded, MNHN- ENSIF2433 ) .
ETYMOLOGY. — Species named after the type locality, plateau of Tankara, near the village of Butmas, Espiritu Santo Is., Vanuatu.
DISTRIBUTION. — Vanuatu, Espiritu Santo Is., inland.
DIAGNOSIS. — Species close to Cardiodactylus cheesmani but differing by colouration darker. Size average, larger than C. enkraussi , shape more stocky than C. novaeguineae ( Figs 8 View FIG ; 17E View FIG ). Male FW generally without a clear light crescent including posterior part of mirror; male genitalia close to that of C. cheesmani , but differing by the shape of the pseudepiphallic sclerite (dorsal crest asymmetrical, basal protuberance near median shrinkage).
DESCRIPTION
Size average, stocky shape. General colouration dark brown, a few specimens lighter, with contrasted yellow or whitish patterns on dorsum and face. Scapes whitish with two dark brown spots, on inner sides and face; antennae homogeneously brown basally and including light areas. Head dorsum with 3 wide dark brown bands, bordered by a thin band behind eye and a yellow line more externally. Fastigium twice as long as wide, dark brown posteriorly, its apex yellow around dark median ocellus. Face with two more or less distinct dark brown curved patterns diverging from front head to below antennae. Mouth parts yellowish, separated from the face by a black line, highlighted by a horizontal yellow band. Palpi whitish, apex darker. Pronotum: dorsal disk trapezoidal, slightly bisinuated posteriorly; dark brown mottled with grey brown and sometimes with whitish, but generally little contrasted. Lateral lobes dark brown to grey brown near dorsal angle; ventral edge whitish. Legs yellowish brown, femora more or less mottled with brown; tibiae more or less banded. Fore tibiae not widened at tympana level. Hind tibiae dorsal face with 6-8 inner (m = 7, n = 8) and 12-15 outer (m =14, n =8) spines above spurs, and 5 or 6 inner (m = 6, n = 8) and 9 or 10 outer (m = 10, n = 8) spines between spurs. Tarsomeres III-1 with 2-6 spines on dorso-external edges (m = 3, n = 8). Hind wings longer than FWs, but proportionally shorter than in C. enkraussi , forming a dark brown tail 1.5 times as long as pronotum. Cerci dark brown, banded but little contrasted.
Male
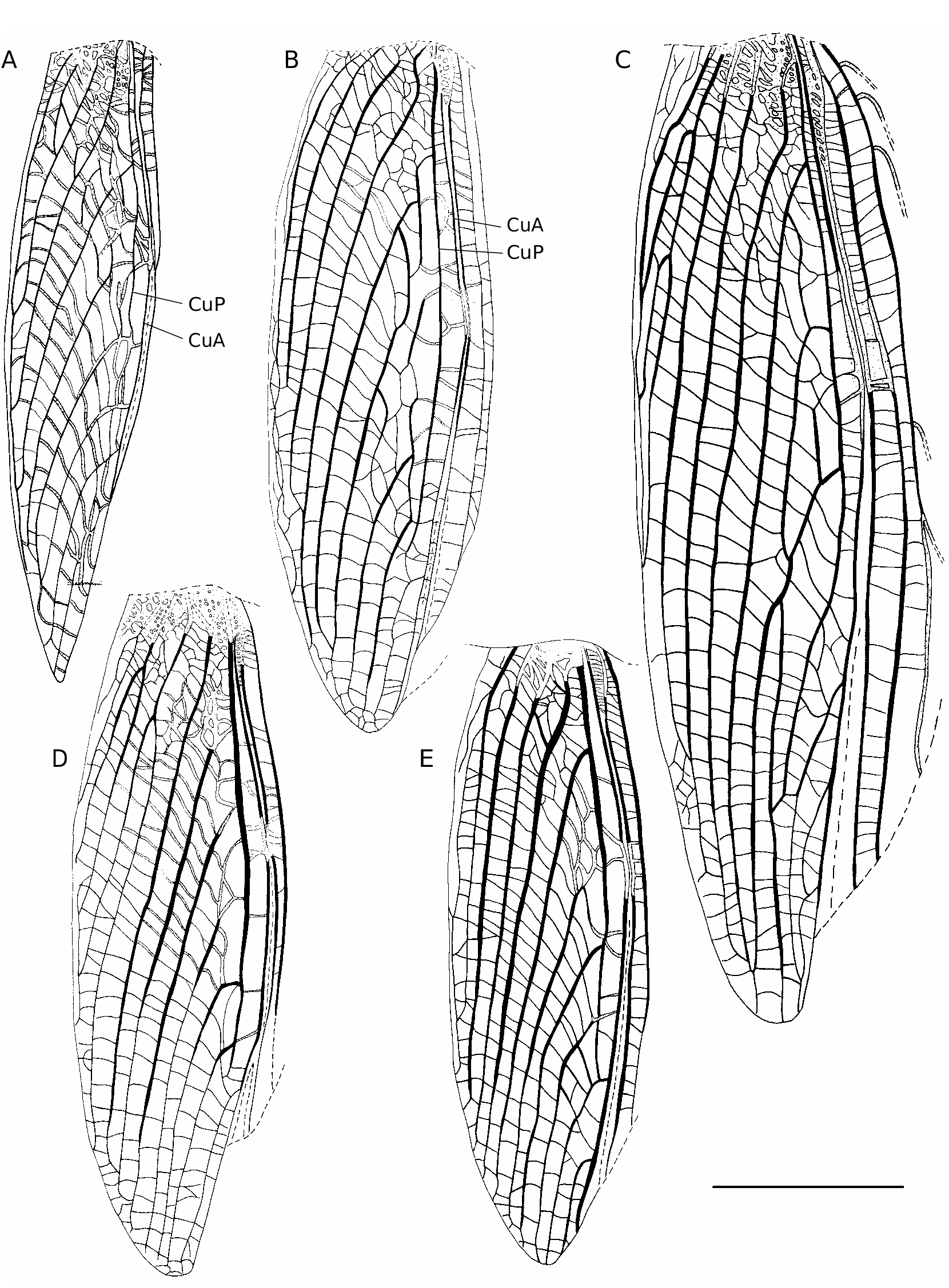
FW colouration ( Figs 1C, D View FIG ; 17E View FIG ): dorsal field generally dark brown, sometimes rust brown; not translucent; veins and cells of similar colour; with constant yellow and/or whitish patterns, including: FW base; 3A and part of 4A; 1A near file angle; harp veins; parts of chords; whitish sclerotized area at harp posterior angle. Harp with a dark triangle behind second harp vein, not distinct in darkest specimens. Transverse pale crescent behind mirror either absent or slightly visible, unlike in C. cheesmani where it is very contrasted and includes part of the mirror ( Otte 2007a: fig. 31). Lateral field dark brown dorsally, including MA, MP, R and bases of veins bifurcating from R; translucent ventrally with whitish veins. FW venation:1A slightly bisinuated;stridulatory vein ( Fig. 3A View FIG ) with 134-166 teeth (m = 156, n = 5), located on both transverse (123-141 teeth, m=134) and basal longitudinal (11-32 teeth, m = 25) parts of 1A. CuP missing.Harp wider than in C. enkraussi , 2W-shaped harp veins; posterior margin raised along diagonal vein. Mirror area variable, c1 long and narrow, c2 quite large; mirror (d1) longer than wide, not rounded, its shape prolonged by d2 and separated at second third by one or several accessory veins. Apical field short, triangular, CuA with 2 or 3 bifurcations posterior to mirror. Lateral field ( Fig. 1D View FIG ): latero-dorsal angle made by MP. R with 8 bifurcating veins.
Male genitalia ( Fig. 4 View FIG F-H): close to C. cheesmani and C. guttulus (Matsumura, 1913) (Robillard & Ichikawa in press). Pseudepiphallus very sclerotized and setose, slightly narrowed at midlength with a basal protuberance absent in C. cheesmani .Two parallel dorsal ridges, forming a gutter, their dorsal edges asymmetrically folded externally, the left one being shorter and widened in all examined males (n = 7; Fig. 4G View FIG ). Pseudepiphallic sclerite extended laterally anterior to the ridges, but without a membranous sac as in C. novaeguineae . Apex of pseudepiphallus spoon-like and rounded, not separate from the rest of pseudepiphallus, its profile differing from C.cheesmani .Rami large,their base at level of pseudepiphallus narrowed part. Pseudepiphallic parameres trilobate, large and close together. Membrane at base of parameres sclerotized. Ectophallus almost hidden within the pseudepiphallus.Ectophallic arc complete, anterior to parameres. Ectophallic apodemes almost reaching rami apex. Apex of ectophallic fold almost hidden by pseudepiphallic parameres, trilobate, the median lobe sponge-like, possibly glandular, shorter than lateral lobes.Endophallic sclerite small, without a medio-posterior expansion. Endophallic apodeme with both lateral lamellas and a short medio-dorsal crest. Membrane of endophallic cavity finely and transversally plicate.
Female
FW colouration: veins and cells dark brown, except: whitish FW base; large whitish spot at first third of length, including CuA and MP; and whitish transverse veins between CuP and CuA. Lateral field as in male. FW venation ( Fig. 5B View FIG ): 9 strong longitudinal veins on dorsal field (including 1-3A). Ovipositor: Shorter than hind femora; apex with both dorsal and ventral edges denticulate, more pronounced on ventral valve ( Fig. 6B View FIG ).
Female genitalia ( Fig. 7 View FIG J-L): copulatory papilla rounded with a basal sclerotized ring; apex narrowed and folded ventrally; ventral preapical convergent crests, concave posteriorly.
Juvenile
First instars observed in laboratory similar to that of C. novaeguineae , yellow with brown longitudinal bands. Later instars with more contrasted colouration, dark brown to black highly mottled with whitish and yellow. Head colouration as in adults, the head dorsum and face sometimes entirely black. Pronotum as in adults, sometimes more contrasted. Metanotum and tergites with median, paired black patterns; rest of abdomen dorsum brown with dark brown dots disposed in longitudinal lines.
Measurements
See Table 7.
HABITAT AND LIFE HISTORY TRAITS
Cardiodactylus tankara n. sp. lives in dense forested areas but is also found near borders and in open areas ( Fig. 17D, E View FIG ). Individuals are found on large leaves of trees, bushes of fern and creepers, or on tree trunks (including Pandanaceae ) at about 2 m height. Singing males are distributed at relatively large distance from one another ( Tables 1; 2). Cardiodactylus tankara n. sp. is sympatric with C. enkraussi at the plateau of Tankara. The population density is comparable to that of C. novaeguineae , and contrasts from the denser populations of C. enkraussi .
BEHAVIOUR
Mating behaviour not observed.
Calling song ( Fig. 15 View FIG ; Table 8)
Males of C. tankara n. sp. are mostly acoustically active at night, but have been heard singing during daytime. The calling song is more conspicuous and noisy than that of sympatric C. enkraussi . Six males recorded in the field at 20-26°C (MNHN-ENSIF 2409, 2411, 2412, 2418, 2419, 2421) are analysed here. At 25°C, the calling song consists in an irregular succession of short echemes (mean echeme duration ± SD = 162 ± 7 ms; period = 2538 ± 1118 ms; duty cycle = 6.9 ± 0.7%; echeme rate = 26.3 echeme/min). The echeme is slightly accelerated from start to end (regular decrease of syllable period within echeme) and shows a crescent amplitude profile. Each echeme is made of 27.5 ± 2.1 syllables with the following characteristics: syllable rate = 170.8 ± 2.1/s; syllable duration = 1.2 ± 0.4 ms; syllable period = 6.2 ± 3.6 ms; syllable duty cycle = 19.5%; and each echeme is finished by a longer syllable, which is likely the contraction of 3 or 4 syllables (mean duration = 4.8 ± 1.9 ms).
The calling frequency of C. tankara n. sp. is relatively high (14.3 ± 0.4 kHz), and corresponds to the third harmonic of the song (Robillard et al. 2007); the mean fundamental frequency is 5.6 ± 0.2 kHz.
Courting song
This song was observed during forced interactions with females of C. novaeguineae ; it is less intense than the calling song, but the echeme is similar to the calling song, except for its irregular period.
Aggressive song
This song was observed in laboratory during interactions between two singing males of C. tankara n. sp. After alternating their songs, one male sometimes charges the other one and sings more frequently in front of him.Th is aggressive song is quite similar to the calling song but of shorter duration and period.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |