
Magelona obockensis Gravier, 1905
|
publication ID |
https://doi.org/ 10.5281/zenodo.198268 |
|
DOI |
https://doi.org/10.5281/zenodo.6208084 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDDD14-E46C-552A-FF41-F958BE21F83E |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona obockensis Gravier, 1905 |
| status |
|
Magelona obockensis Gravier, 1905 View in CoL
Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
Magelona obockensis Gravier, 1905: 44 View in CoL –45; 1906: 163–166, pl. 2, figs 186–192 Magelona heteropoda Mohammed, 1973 View in CoL (see below)
Magelona papillicornis View in CoL (not F. Müller, 1858)— Amoureux 1983; 738, fig. 7 (see below)
Material examined. Obock, Djibouti, Gulf of Arden, Red Sea—in the sands of Balanoglossus with Cymodoce , low tide, 7 syntypes ( MNHN Type 1357; 7 af, 1 pf, 9 f), 5th March 1904 (note Gravier examined 12 specimens for the original description, the largest being 66 mm in length for 131 chaetigers, however this specimen does not appear to be present in the syntype material, and is somewhat larger than the other syntypes).—Locality and collector information unknown ( MNHN A 172) from general collection; 1 c (posteriorly regenerating), 1 af (heavily dissected).
Diagnosis. Prostomium longer than wide, slender triangular, anteriorly rounded without prostomial horns. Notopodia of chaetigers 1–8 with large foliaceous postchaetal lamellae, upper edges minutely crenulate, bilobed on several chaetigers. Slender cirriform dorsal processes on all thoracic chaetigers. Neuropodia with slender cirriform ventral processes that become shorter and broader along thorax. Neuropodia of chaetiger 8 with distinct digitiform prechaetal processes, in addition to subtriangular postchaetal lamellae. Chaetiger 9 with broadly rounded low postchaetal lamellae, and small papilliform processes present at the inner margins of chaetal rows in both the noto– & neuropodia. Chaetigers 1–8 with capillary chaetae, chaetiger 9 with mucronate chaetae. Abdominal lateral lamellae spatulate, bluntly pointed, basally constricted. Hooded hooks tridentate, in two groups, vis–à–vis. Anteriorly and posteriorly open pouches present on abdominal chaetigers.
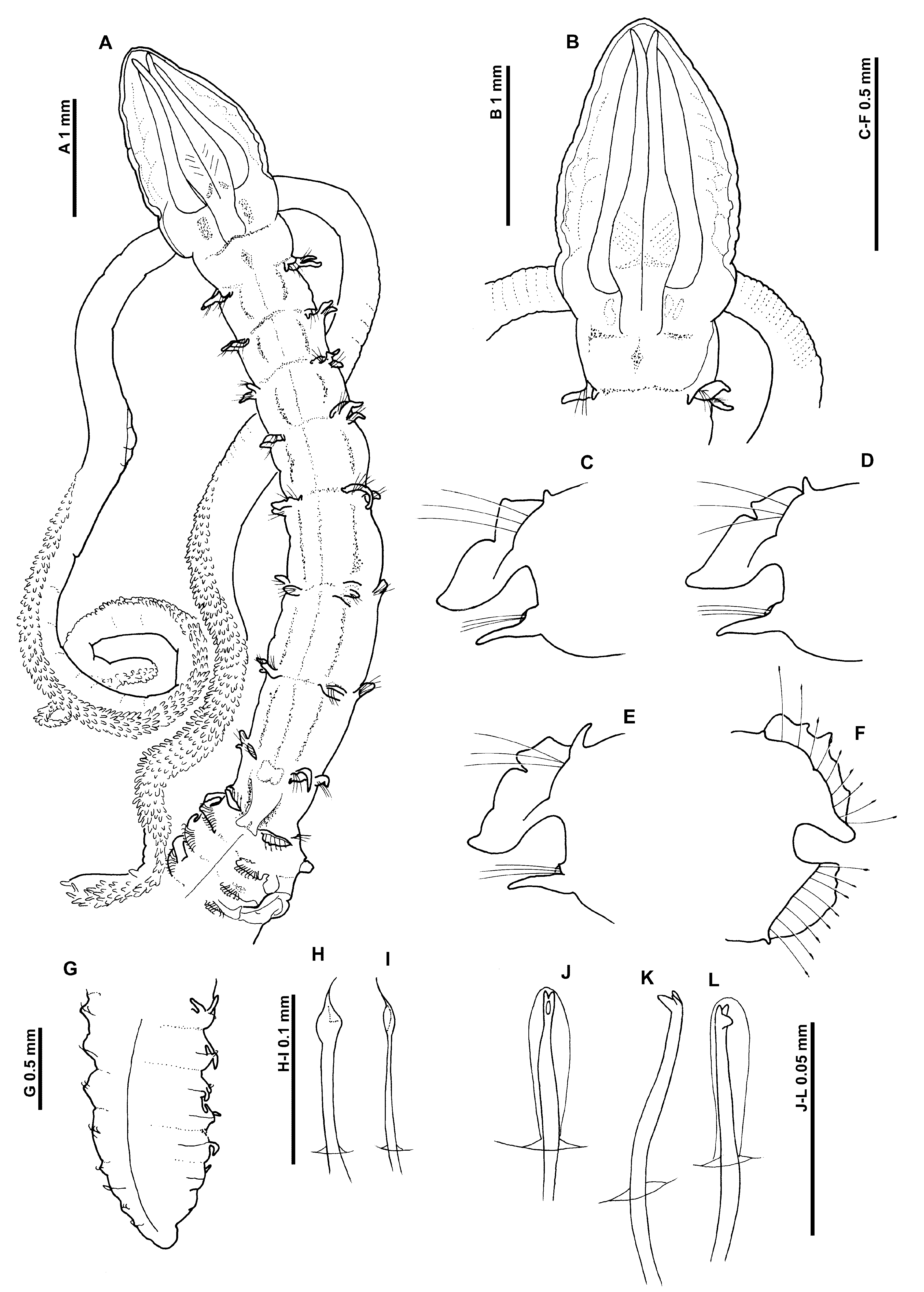
Description. A moderately stout species; abdomen markedly wider than thorax ( Fig. 1 View FIGURE 1 A). Syntypes all incomplete, condition generally poor. Dimensions of broadest specimen: Prostomium 1.87 mm long, 1.15 mm wide; thorax (including prostomium) 7.05 mm long, 0.83 mm wide; abdomen 1.1 mm wide; total length 19.85 mm for 39 chaetigers. Longest specimen, 22.1 mm for 53 chaetigers. Other syntypes 11.4 mm – 17.2 mm with 22–36 chaetigers. Complete specimen (MNHN A 172) ~ 31 mm for 81 chaetigers (4–5 posteriormost chaetigers regenerating), with marked difference between thorax and abdomen, constricted at chaetiger 9.
Prostomium elongate, much longer than wide (L:W ratio 1.4–1.88), slender triangular, without prostomial horns, anterior margin smooth, rounded (sometimes formed into a blunt tip); lateral edges minutely wavy; eyes absent. Two pairs of prominent longitudinal dorsal muscular ridges, outer pair (slightly shorter) abutting inners for entire length, inner pair reaching the distal tip of prostomium, where they diverge only very slightly ( Fig. 1 View FIGURE 1 B). Indistinct quadrangular areas (poorly preserved) either side of muscular ridges, distinct in general collection specimens (MNHN A 172). Proboscis everted in four specimens, heart–shaped when fully everted, oval when partially everted; inferiorly longitudinal ridged, appearing smooth superiorly but with light transverse ridging. Palps arising ventrolaterally from base of prostomium, robust, tapering to fine translucent tips in figured specimen ( Fig. 1 View FIGURE 1 A) (although not observed in other specimens), reaching at least chaetigers 14– 17, non–papillated region reaching chaetigers 2–3. Papillae very short proximally, increasing gradually in size, papillae long at distal tip. Initially 3–4 rows of papillae either side of inconspicuous ventral groove, distally 2 rows. Palps of complete specimen (MNHN A 172) more slender, although still robust, with 1–2 papillae either side of groove, reaching about chaetiger 12 (possibly regenerating?).
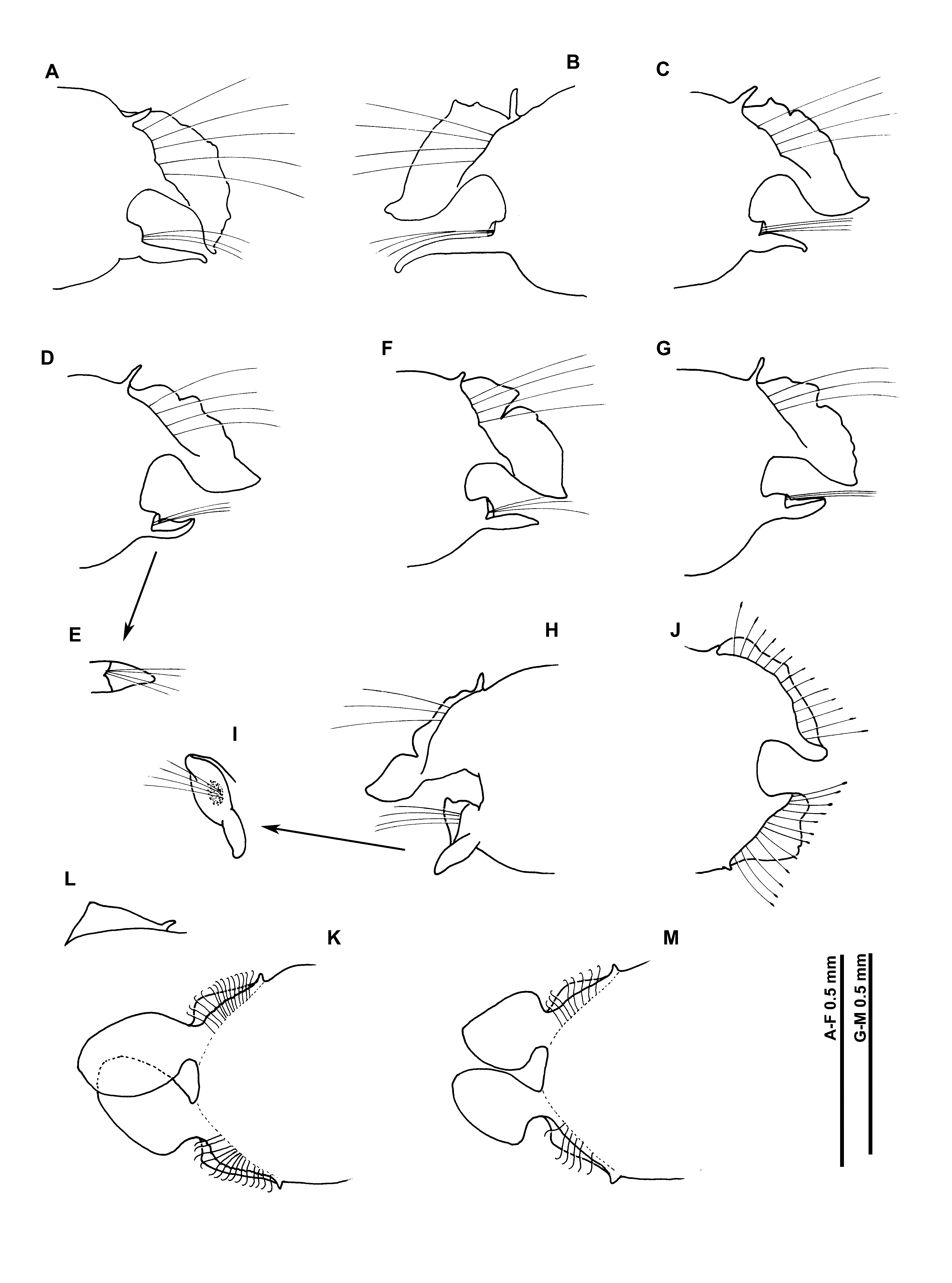
Peristomium achaetous, roughly twice the size of chaetiger 1. Chaetigers 1–7 similar; parapodia biramous. Notopodia with well–developed, thick, low prechaetal lamellae confluent with large delicate foliaceous postchaetal lamellae, latter with pointed tips, of similar size throughout thorax ( Figs 1 View FIGURE 1 C–E, 2A–G). Upper edges of notopodial postchaetal lamellae crenulated, degree of crenulation variable between specimens, with a discrete notch usually present by the fourth or fifth chaetigers ( Figs 2 View FIGURE 2 C–D), developing into distinct bilobed lamellae from at least chaetiger 6 ( Figs 2 View FIGURE 2 F–G). Most notopodial thoracic lamellae of syntype material either damaged or missing (due to delicate nature of the postchaetal lamellae, particularly when bilobed), thus position of first appearance of bilobed lamellae often impossible to state. However, broadest syntype with bilobed lamellae from chaetiger 1 ( Figs 1 View FIGURE 1 C–E), although not present until posterior thoracic chaetigers in other specimens. Single long, slender, tapering cirriform prechaetal superior process (DML) present on notopodia of all thoracic chaetigers, gradually increasing in size to chaetiger 5 but decreasing towards the posterior thorax.
Neuropodia of chaetigers 1–7 with single long slender cirriform processes ventrally (VNL), directly under chaetae but becoming distinctly prechaetal by chaetiger 7, distal tips pointed. Longest on chaetiger 2, becoming shorter and broader, towards posterior thorax. Pre– and postchaetal lamellae well–developed, encircling chaetae cuff–like.
Chaetiger 8 ( Fig. 2 View FIGURE 2 H): Notopodial lamellae noticeably bilobed, distal portion subtriangular, prechaetal superior processes small (DML). Neuropodial processes prechaetal, rounded and digitiform. Low prechaetal lamellae confluent with subtriangular postchaetal lamellae, encircling chaetae superiorly, cuff–like ( Fig. 2 View FIGURE 2 I). Chaetae of chaetigers 1–8 simple winged capillaries.
Chaetiger 9 ( Fig. 1 View FIGURE 1 A): shorter and narrower than preceding chaetigers. Notopodial prechaetal lamellae low, minutely crenulated and broadly rounded, confluent with higher postchaetal lamellae ( Figs 1 View FIGURE 1 F, 2J) (crenulate/pectinate in some specimens, degree of crenulation often varying between notopodia on both sides of same example); lamellae encircling chaetae underneath as lateral expansions. Superior processes (DML) very small to minute, obscured/missing(?) on several specimens. Neuropodia similar to notopodia, without lateral expansions, small ventral prechaetal processes present (VML of some authors). Chaetae spatulate ( Fig. 1 View FIGURE 1 H), arranged in arcs; chaetae longer but with narrower distal tips towards margins of each fan ( Fig. 1 View FIGURE 1 I).
A conspicuous, oval swelling present ventrally on all specimens, level with the lamellae of chaetiger 8 ( Fig. 3 View FIGURE 3 ). Additional ventral swellings observed on earlier chaetigers, as two oblique anterior facing structures (often reniform) ( Fig. 3 View FIGURE 3 ), initially poorly developed at chaetiger 1, increasing in development along thorax. Those of chaetiger 7 show some variation in size/shape, sometimes similar to preceding chaetigers or more well–developed, almost abutting. Swellings with sporadic white speckles. An occasional oval dorsal swelling, observed ( Fig. 1 View FIGURE 1 A) between parapodia of chaetiger 8.
Abdominal chaetigers ( Figs 2 View FIGURE 2 K–M) with broad, spatulate lateral lamellae, of about equal size in both rami, basally constricted, bluntly pointed. Lamellar shape shows some variation (even within space of several segments along a specimen, and between rami of same parapodium); from rounded, subrectangular to slightly reniform. Postchaetal extension of lateral lamellae, behind chaetal rows well–developed, especially in anterior abdomen, apex forming a distinct rounded tip in some specimens ( Fig. 2 View FIGURE 2 L), similar to that figured by Gravier (1906: fig. 331). Small triangular processes (DML & VML) present at inner margins of chaetal rows.
Lamellae of posterior fragment ( Fig. 1 View FIGURE 1 G) (syntype, 21 chaetiger pf) much reduced, initially still fairly broad and basally constricted around 20 chaetigers from pygidium, reducing to cirriform.
Abdominal chaetae ( Figs 1 View FIGURE 1 J–L) tridentate hooded hooks of similar size, superior two fangs parallel, above main fang. Hooks in two groups in each ramus, main fangs vis–à–vis; group at the inner margins of chaetal rows with around twice number of chaetae ( Fig. 2 View FIGURE 2 K). Initially around 10–14 hooks per rami, decreasing to approximately 8–10 hooks per rami on posterior fragment.
Paired anteriorly open pouches between chaetigers 11 and 12 (Σ configuration of Fiege et al. 2000 —see discussion), well–developed, large membrane, often extruded, bounded between two cuticular flaps. Unpaired posteriorly open pouches (C configuration of Fiege et al. 2000 —see discussion) present in posterior abdomen, generally on alternate chaetigers. Pouches differing from previously described C configuration pouches, often quite large, more expanded both dorsally and ventrally, often convoluted. On longest observed syntype (53 chaetiger af) pouches observed on 11, 16L, 18R, 20L, 21R, 23L, 25R, 27L, 29R, 31L, 32R, 34L, 36R, 38L, 40R, 43L, 44R, 45L, 47R, 49L, 50L, 51R, 52L; exact position of pouches often difficult to discern due to condition of material. On this specimen, unpaired anteriorly open pouches observed on chaetigers 16L, 18R, 20L, 21R, 25R, differing from the paired pouches seen between chaetigers 11 and 12 and more closely resembling the posteriorly open pouches of later chaetigers. On other syntype material where pouches could be observed, all pouches after the initial pair, open posteriorly. Posteriorly open pouches observed on posterior fragment (14th & 17th chaetiger from pygidium). No anal cirri observed on posterior fragment ( Fig. 1 View FIGURE 1 G), although Gravier originally described two short sub–terminal anal cirri as present.
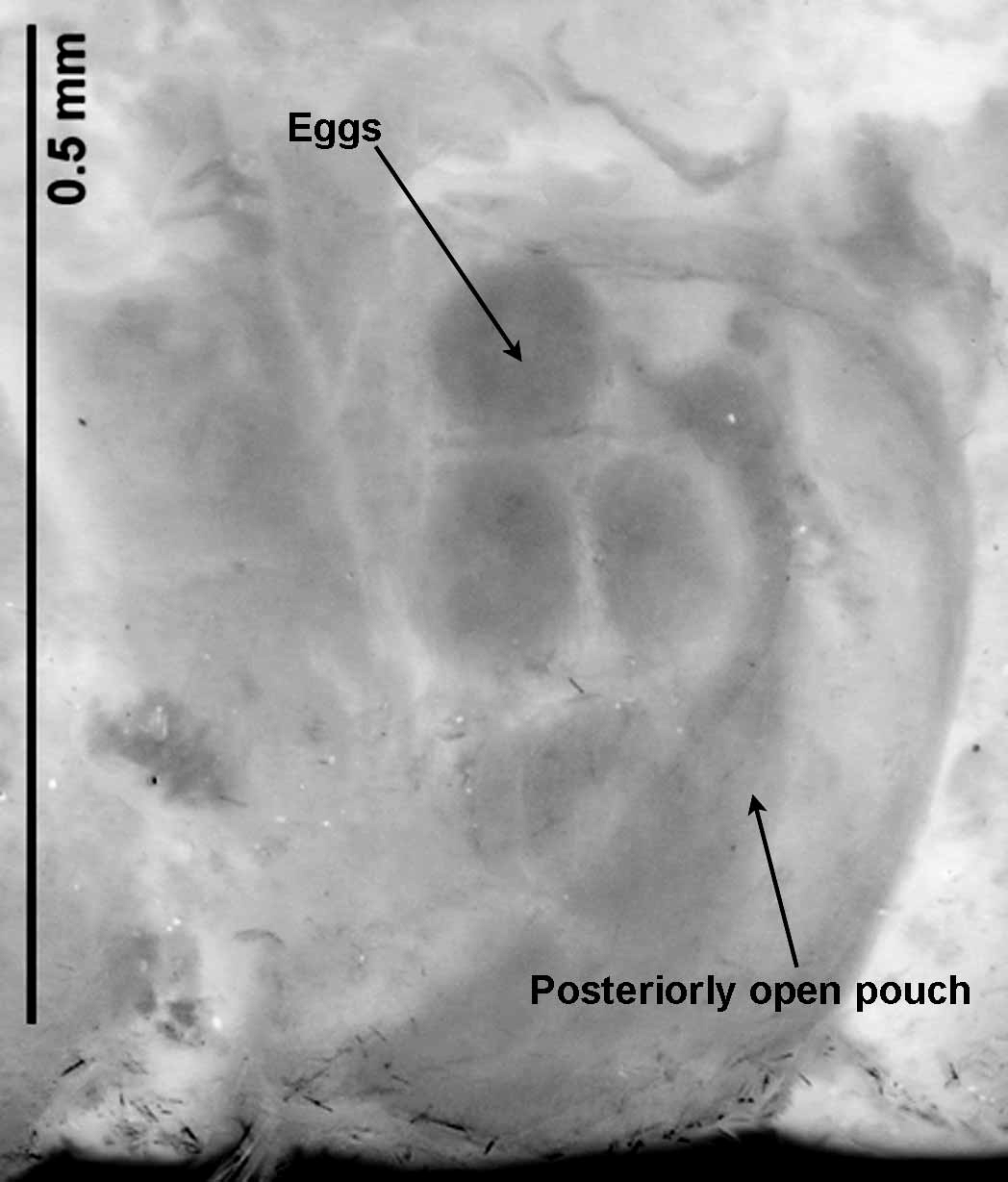
Several fragments (1.8 mm fragment for 4 chaetigers, 14 mm (aprox.) fragment for 40 chaetigers) ovigerous, eggs appearing to be compartmentalized and associated with ‘pouches’ ( Fig. 4 View FIGURE 4 ). These structures are similar to posteriorly open pouches as seen on anterior fragments, and there appears on several to be an ‘opening’. However, the poor condition of these fragments makes it difficult to ascertain whether these are pouches, or whether they are simply ‘swellings’ of the body wall where the epidermis has split. The larger of the two fragments also contains several different sizes of eggs. Due to the condition of the specimens, nothing more can be interpreted from them. No eggs observed in anterior fragments.
Colour. The colouration originally described by Gravier was pale pink in the anterior, with a brownish posterior. He noted brown pigmentation on the palps, which increased towards the distal tip, and dark tinted band outside and in immediate contact with the external row of papillae. This pigmentation has been lost in the preserved material, which is now generally pale, with light brown/yellow colouration. Staining with methyl green shows no clear pattern, but an overall diffuse stain. However, after staining, when much of the stain has dissipated, green speckled areas associated with the ventral swellings can be observed as well as, a parallel longitudinal stripe either side of the mid–dorsal line in thorax (at the level of the superior processes), strong particularly between chaetigers 1–4. Some staining evident abdominally between the parapodia.
Habitat. Type specimens found in sand, at low tide at Obock. Also recorded in sand, intertidally from the Red Sea: Na’ma Bay, Israel ( Amoureux 1983: 737–738, Table 1, fig. 7, specimens unavailable for verification). Other specimens identified as M. papillicornis in the same study have been shown to be M. obockensis , see below).
Remarks. Several features observed differ from those of the original description of this species, in particular, the bilobed nature of the lamellae of the thorax. Gravier (1906: fig. 327) described and drew the 7th chaetiger, however, both the description and drawings differ from the observed syntype material. The bilobed lamellae are particularly delicate, and are prone to splitting along the junction between the two lamellar regions. As a consequence, many of the specimens have lost the distal portion of these lamellae, making a complete set of drawings from one specimen difficult. It is entirely possible that his figure was drawn from a parapodium in which the lamella was incomplete, or drawn from a parapodium in the anterior thorax. Gravier also stated that dorsal cirri, above the lamellae, were not present on the 1st or the 9th chaetigers. Small superior prechaetal processes (DML) have been observed to be present on these chaetigers within the syntype material. However, these are small, especially on chaetiger 9, and maybe obscured or sometimes missing?
Magelona obockensis View in CoL belongs to the ‘ Magelona mirabilis View in CoL group’ of species, all having a prostomium lacking frontal horns, and possessing spatulate chaetae on the chaetiger 9. Members of this group are: M. mirabilis ( Johnston, 1865) View in CoL , M. johnstoni Fiege, Licher & Mackie, 2000 View in CoL , M. crenulata Bolívar & Lana, 1986 View in CoL , M. pitelkai Hartman, 1944 View in CoL , M. sacculata Hartman, 1961 View in CoL , M. riojai Jones, 1963 View in CoL , M. conversa Mortimer & Mackie, 2003 View in CoL , M. pectinata Nateewathana & Hylleberg, 1991 View in CoL , M. sachalinensis Buzhinskaja, 1985 View in CoL , and two unnamed species (M. sp. A and M. sp. B) from the Gulf of Mexico ( Uebelacker & Jones 1984). A further two species, M. heteropoda View in CoL and M. tinae View in CoL will be discussed further (see below).
Magelona obockensis View in CoL differs from M. conversa View in CoL , M. sachalinensis View in CoL and M. sp. A in possessing tridentate and not bidentate hooded hooks. It differs from M. pectinata View in CoL , M. riojai View in CoL , M. sacculata View in CoL and M. mirabilis View in CoL in possessing hooded hooks in two groups vis–à–vis, and not one unidirectional facing group. This is assumed from the drawings in M. pectinata View in CoL as it was not stated, however M. pectinata View in CoL further differs in possessing pectinate processes on the upper edges of the thoracic postchaetal lamellae rather than crenulated. It differs from M. pitelkai View in CoL (see Jones 1978) and M. johnstoni View in CoL in possessing bilobed notopodial lamellae of the posterior thorax. Magelona obockensis View in CoL shares similarities with M. crenulata View in CoL and M. sp. B in the shape of the thoracic lamellae (see Bolívar & Lana 1986, fig. 44; Uebelacker & Jones 1984, fig. 7–4). However, both differ in possessing both postchaetal and ventral neuropodial lamellae in the thorax.
Magelona papillicornis View in CoL , originally described from Brazil was reported as being present in the Red Sea by Amoureux (1983). A single specimen of M. papillicornis View in CoL (MNHN A895) labelled Na’ama Bay (22nd July 1979) Sinaï Golfe d’Aqaba, was borrowed from the Muséum National d'Histoire Naturelle, Paris. Amoureux described finding two specimens of M. papillicornis View in CoL from Na’ama Bay, although only one specimen was present in this museum vial. However, the location details of this specimen agree well with those recorded, and is herein considered as that described by Amoureux. Detailed examination of this specimen shows that it is actually M. obockensis View in CoL and shows no similarity with M. papillicornis F. Müller View in CoL as redescribed by Jones (1977). Unfortunately, the specimens which Amoureux identified as M. obockensis View in CoL could not be located for cross–comparison.

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Magelona obockensis Gravier, 1905
| Mortimer, Kate 2010 |
Magelona obockensis
| Gravier 1905: 44 |