
Chactopsis carolinae, Botero-Trujillo, Ricardo, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181581 |
|
DOI |
https://doi.org/10.5281/zenodo.5678592 |
|
persistent identifier |
https://treatment.plazi.org/id/03BD87E9-FFD7-FF8E-4BB9-2E21FC8870C2 |
|
treatment provided by |
Plazi |
|
scientific name |
Chactopsis carolinae |
| status |
sp. nov. |
Chactopsis carolinae sp. nov.
( Figures 1–14 View FIGURE 1 View FIGURES 2 – 5 View FIGURES 6 – 14 , Table 1 View TABLE 1 )
Type material.— COLOMBIA. Male holotype (adult) from Vichada department, Cumaribo, Selva de Matavén, 04°29’13’’ N, - 68°00’22’’ W, 260m, Bosque de Planicies Arenosas, Pitfall, 31/03/2007 – 02/04/2007, L. E. Franco (IAvH-E 100759). One female paratype (adult) from Vichada department, Cumaribo, Selva de Matavén, 04°36’33’’ N, - 67°51’52’’ W, 300m, Bosque de Cerro Rocoso, Winkler, 06–08/03/2007, L. E. Franco (IAvH-E 100760). One female paratype (juvenile) from Vichada department, Cumaribo, Selva de Matavén, 04°33’32’’ N, - 68°11’51’’ W, 270m, Bosque de Tierra Firme, Pitfall, 22–24/03/2007, L. E. Franco (IAvH-E 100761).
Etymology.— Patronym dedicated to the author’s girlfriend, Biviana Carolina Barón Mendoza , in acknowledgement to her support, encouragement and inspiration. It celebrates both past and forthcoming experiences together.
Diagnosis.— Chactopsis carolinae sp. nov. is most similar to Ch. anduzei , with which it shares the following combination of features that distinguishes both species from all other in the genus: the trichobothrium ib is not located in a basal position on the fixed finger of pedipalp chela ( Fig. 9 View FIGURES 6 – 14 ; González-Sponga 1982: Fig. 2 View FIGURES 2 – 5 ); eb and esb are located between Db and Dt ( Figs. 6–7 View FIGURES 6 – 14 ; González-Sponga 1982: Figs. 1–2 View FIGURE 1 View FIGURES 2 – 5 ); pedipalp patella bears 23 trichobothria on the external surface with esb5 absent ( Fig. 13 View FIGURES 6 – 14 ; González-Sponga 1982: Fig. 3 View FIGURES 2 – 5 ), and seven ventral trichobothria of which v6 and v7 are not arranged linearly with v1-v 5 in the ventroexternal border but located near to the median longitudinal line of the segment ( Fig. 12 View FIGURES 6 – 14 ; González-Sponga 1982: Fig. 4 View FIGURES 2 – 5 ). Furthermore, it trichobothrium is located between eb and esb, and ib is considerably basal to eb ( Figs. 6–7 View FIGURES 6 – 14 )— although González-Sponga (1982: 128) erroneously noted that in Ch. anduzei ib and it trichobothria are located between eb and esb (see González-Sponga 1982: Fig. 2 View FIGURES 2 – 5 ). The new species can be readily distinguished from Ch. anduzei since in the former the trichobothrium db is basal in relation to Et4 ( Fig. 7 View FIGURES 6 – 14 ), patellar em2 is dorsal to est5 ( Fig. 13 View FIGURES 6 – 14 ), sternite VII lacks any vestige of paramedian carinae, and metasomal segment V bears well-defined dorsolateral carinae. In contrast, in Ch. anduzei the trichobothrium db is distal in relation to Et4 ( González-Sponga 1982: Fig. 1 View FIGURE 1 ; Lourenço & Francke, 1986), patellar em2 is ventral to est5 ( González-Sponga 1982: Fig. 3 View FIGURES 2 – 5 ), sternite VII presents paramedian carinae formed by vestigial granules, and metasomal segment V does not have well-defined dorsolateral carinae ( González-Sponga 1982). In addition, the coloration pattern is different, especially on the tergites where a broad longitudinal yellowish band is present in the new species ( Figs. 2, 4 View FIGURES 2 – 5 ) and absent in Ch. anduzei ( González-Sponga 1982) .
Description of the male holotype (IAvH-E 100759).
Coloration.— Carapace predominantly brown with some lateral yellowish regions and a broad median longitudinal yellow area crossing from the anterior to the posterior border; median and lateral ocular tubercles black; a brown area is present from the median ocular tubercle to the anterior margin of the carapace, this is more conspicuous in the posterior region and is formed by inconspicuous spots anteriorly. Chelicerae predominantly yellow; coxa with some light-brown regions in dorsal view; tibia dorsally and externally with broad brown longitudinal reticulations and others thinner located between these, a rounded brown area is present just under the base of the fixed finger, ventral and internal surfaces with inconspicuous reticulations; fixed finger yellow, teeth reddish; movable finger light-brown, teeth reddish. Coxosternal region almost completely yellow, except for coxae II that bear few brownish spots in the inner half of the segment, and the sternum that is brownish anteriorly. Genital opercula, pectinal basal piece, pectines and sternites III–IV completely yellow; sternites V–VI with inconspicuous brown spots laterally; sternite VII with dark-brown regions on the position of lateral carinae. Tergites predominantly brown; a broad median longitudinal yellowish band is present in all tergites ( Figs. 2, 4 View FIGURES 2 – 5 ); each side of tergites II–VI with two transverse yellow lines converging near to the longitudinal line, resembling an arrow; lateral margins of all tergites yellow; tergite VII with brown mottling laterally. Metasoma with abundant dark-brown mottling in lateral and ventral surfaces; dorsal region of all segments completely yellow, except for segment V that bears lateral brown-spotted areas posteriorly. Telson yellow in dorsal surface, with few brown spots basally; lateral and ventral surfaces predominantly brown, with two ventromedian and two lateral yellow bands; aculeus reddish and spotless. Pedipalps predominantly light-brown; femur with brown regions in dorsointernal, dorsal, and ventroexternal surfaces, others yellow; patella with yellow areas in dorsal, external and internal surfaces, predominantly yellow ventrally; hand with longitudinal brown bands and few reticulations in all surfaces, except ventrally where almost completely yellow; fixed and movable fingers yellow. Legs with variegated pigmentation, except for coxae and trochanters that are yellow.
Morphology.— Carapace: subtriangular, densely covered with rounded granules throughout; anterior margin moderately emarginated and with a median tip directed anteriorly ( Fig. 14 View FIGURES 6 – 14 ); median ocular tubercle low, distinctly anterior to the centre of the carapace; lateral ocular tubercles each with thee ocelli, the posterior most is reduced ( Fig. 14 View FIGURES 6 – 14 ). Chelicerae: with abundant setae on internal and ventral surfaces; cheliceral dentition characteristic of the family Chactidae ( Vachon 1963) . Coxosternal region: sternum pentagonal, with a median longitudinal furrow; coxae sparsely punctulate, with few marginal weak tubercles and few setae; coxapophyses I–II with dense pilosity anteriorly. Genital opercula and pectines: genital opercula divided longitudinally; pectinal basal piece wider than long, scarcely punctulate; pectines long, surpassing the lateral margins of sternite III and with 11:11 teeth. Sternites: III–VII smooth, densely punctulate; sternite VII with sparse weak granules on the lateral margins, without any vestige of paramedian carinae but with two slight elevations on the position of lateral carinae; spiracles small and oval. Tergites: with similar granulation to that of the carapace; tergites III–VI with vestigial axial and dorsolateral carinae represented by one or two granules on the posterior borders; tergite VII tetracarinate (paired dorsolateral and lateral carinae, incomplete and with strong but non-spinoid granules), a median elevation is present in the position of the axial carina, where some greater granules are located in unaligned position. Metasoma: with abundant long setae; segments I–IV with sparse and weak granulation throughout; segment V with dense granulation; all carinae vestigial, slightly elevated except for dorsal and dorsolateral carinae that are stronger; segment I with eight complete carinae (paired ventrolateral, intermedian, dorsolateral and dorsal carinae; ventral carinae absent); segment II with ten (ventral carinae incomplete, reduced to a median elevation; intermedian carinae only represented posteriorly by a big tubercle; others complete); segment III with ten (intermedian carinae as in segment II; others complete); segments IV–V with eight (intermedian carinae absent; others complete); in segment V dorsolateral carinae are only present on the anterior and median thirds of the segment, and ventral carinae are diffuse, difficult to distinguish from the remaining granules of the tegument. Telson densely punctulate throughout and with welldeveloped granules in lateral and ventral surfaces; aculeus short, thick basally. Pedipalps: femur tricarinate, with sparse rounded granules in dorsal and internal surfaces; patella tricarinate, with few low granules dorsally; chela acarinate, smooth; all pedipalp segments densely punctulate, except dorsal surfaces of femur and patella. Trichobothriotaxy ( Figs. 6–13 View FIGURES 6 – 14 ): Type C ( Vachon 1973) with 62 trichobothria; femur with three trichobothria; patella with 33 of which 23 are external; chela with 26. Legs: tibia, basitarsus and telotarsus with abundant setae not arranged in rows on external, internal and ventral surfaces.
Comparisons with the adult female paratype (IAvH-E 100760).
The following features differ from the holotype: Coloration.— coxae II, sternites V–VI, and pedipalp patella completely yellow; pedipalp femur predominantly yellow, except for two distal brown areas, one dorsal and the other ventral. Morphology.— Ven t er: pectines with 8:8 teeth (idem in the juvenile female). Metasoma: segment I bears vestigial, slightly elevated ventral carinae.
Distribution and landscape.— This species is known only from the type locality: Selva de Matavén, Cumaribo, Vichada department ( Fig. 1 View FIGURE 1 ). This territory is conformed of alluvial valleys and rocky high plains ( IGAC 1996). The area presents semiarid zones, original forests, savannas both natural and produced through human activities, and relicts of felled forests ( Molano 1998).
Even though the known distribution of Ch. carolinae sp. nov. is relatively close to the type locality of Ch. anduzei (Caño Pava, between El Danto and Gavilán, Atures department, Venezuela), the former inhabits the Negro-Branco Moist Forests ecoregion in Colombia, whereas the second occurs in Venezuela—at least on the basis of González-Sponga’s (1996: 112) map—in the Guayanan Highlands Moist Forests (National Geographic Society 2001). The Negro-Branco Moist Forests ecoregion occurs along the Negro River basin, along the southwestern edge of the Guayana Shield in eastern Colombia, northwestern Brazil, and most of southern Venezuela. It is strongly influenced by numerous rivers and streams that form the Negro River and Branco River watersheds, and is known to be a region of high species endemism (National Geographic Society 2001).
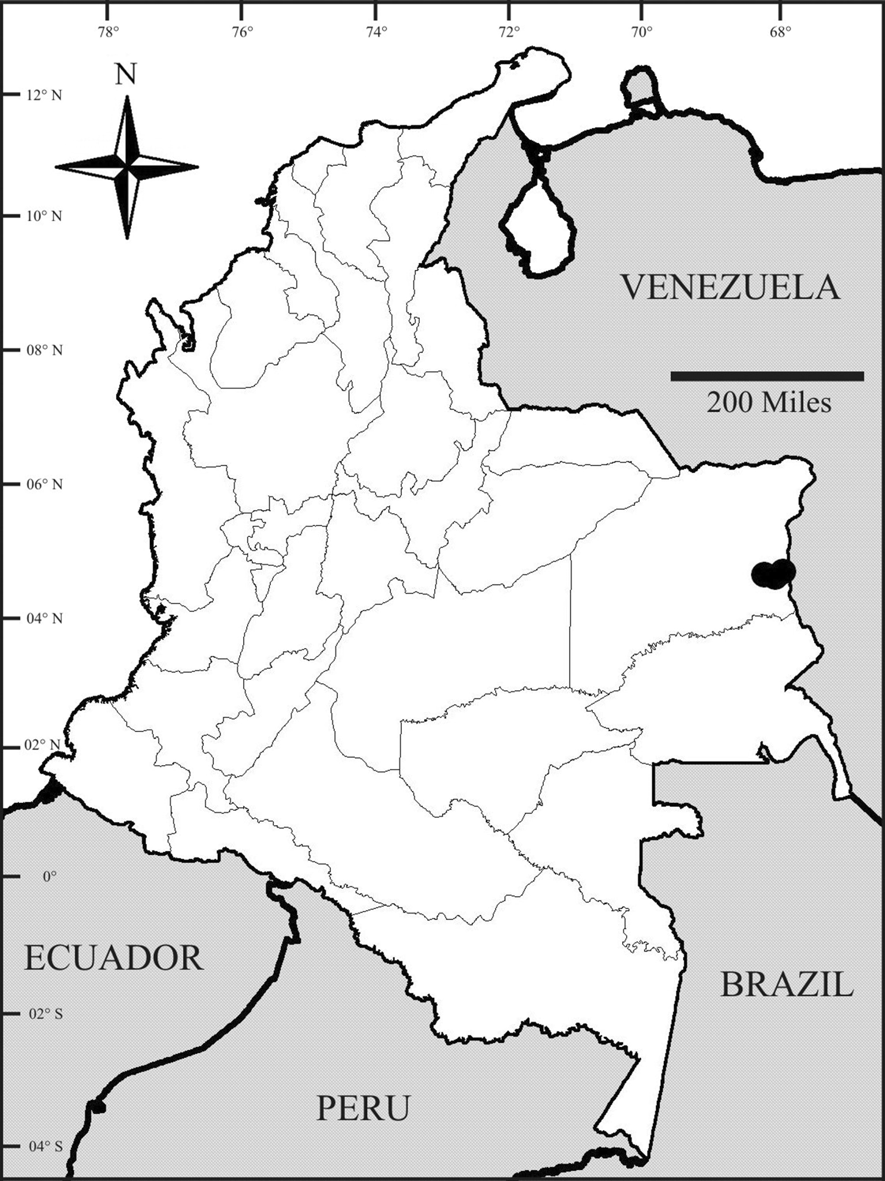

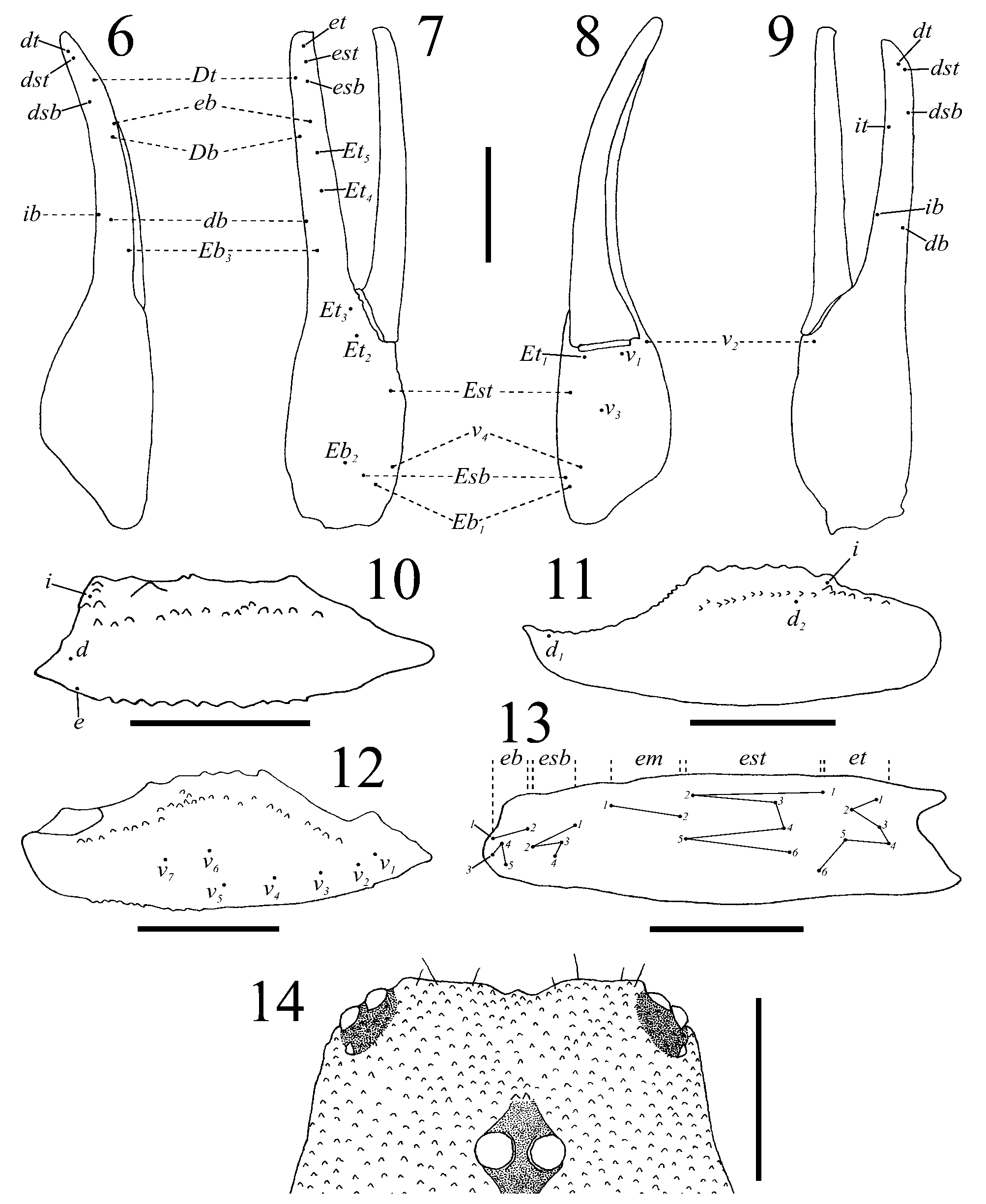
TABLE 1. Morphometric measurements (in mm) of male holotype and adult female paratype of Chactopsis carolinae sp. nov.
| Male holotype (IAvH-E 100759) | Female paratype (IAvH-E 100760) | |
|---|---|---|
| Total body L (including telson) | 18.99 | 22.67 |
| Carapace Length Anterior W | 2.64 1.66 | 3.02 2.11 |
| Posterior W Ocular diameter | 2.52 0.12 | 3.24 0.18 |
| Interocular distance Mesosoma Total L | 0.14 5.08 | 0.16 6.42 |
| Metasoma Total L (including telson) Segment I L | 11.27 0.99 | 13.23 1.23 |
| Segment I W Segment I D | 1.56 1.21 | 2.04 1.39 |
| Segment II L Segment II W | 1.14 1.50 | 1.36 1.95 |
| Segment II D Segment III L | 1.11 1.20 | 1.44 1.44 |
| Segment III W | 1.60 | 1.95 |
| Segment III D Segment IV L Segment IV W | 1.21 1.63 1.60 | 1.62 1.85 1.99 |
| Segment IV D | 1.18 | 1.81 |
| Segment V L Segment V W Segment V D | 3.03 1.60 1.31 | 3.57 2.01 1.74 |
| Telson L Vesicle W | 3.28 1.24 | 3.78 1.52 |
| Vesicle D | 0.94 | 1.19 |
| Pedipalps Total L Femur L Femur W | 8.55 2.10 0.74 | 10.49 2.41 0.83 |
| Patella L | 2.81 | 3.22 |
| Patella W Chela L | 0.99 3.64 | 1.24 4.86 |
| Chela W Chela D Movable finger L | 0.90 0.96 2.25 | 1.18 1.19 3.23 |
| Palm L | 1.53 | 1.78 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.