
Halicyclops continentalis, Ueda & Nagai, 2009
|
publication ID |
https://doi.org/ 10.1080/00222930802585810 |
|
persistent identifier |
https://treatment.plazi.org/id/03BD3F5A-F865-FFDA-C7C2-FB6E949AF932 |
|
treatment provided by |
Carolina |
|
scientific name |
Halicyclops continentalis |
| status |
|
Genus Halicyclops A. M. Norman, 1903
Halicyclops continentalis sp. nov.
( Figures 2 View Figure 2 , 3 View Figure 3 )
Synonym
Halicyclops sinensis, Tai and Chen (1979) , p. 313–315, figures 176, 177.
Type material
One female holotype ( NSMT Cr-18237) dissected and mounted on two glass slides, and eight undissected female paratypes ( NSMT Cr-18238) preserved in alcohol; collected from the Honmyo-gawa River mouth on 28 August 1997 .
Type locality
Honmyo-gawa River mouth (32 ° 539020 N, 130 ° 089100 E) in a detention pond of Isahaya Bay, Kyushu, Japan .
Etymology
The specific epithet refers to the primary distribution of the species along the coast of the East Asian continent.
Description
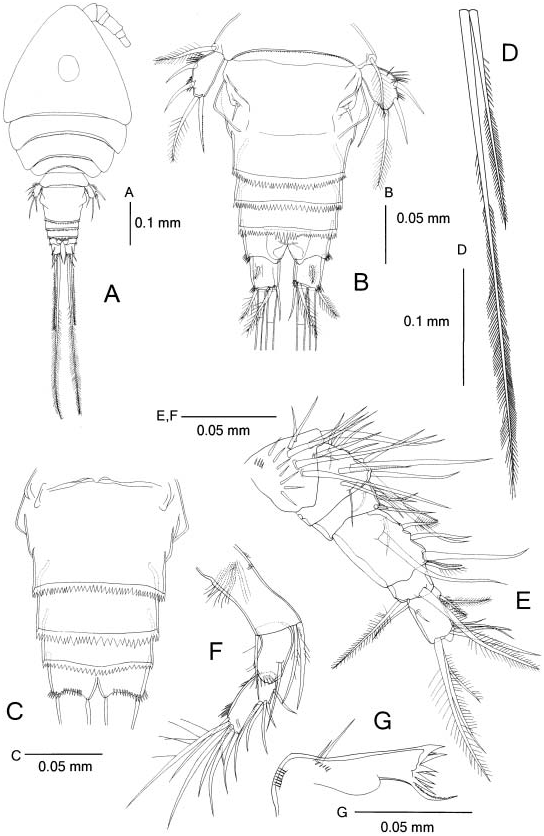
Female. Body ( Figure 2A View Figure 2 ) length 0.55–0.59 mm (n 55, holotype 0.59 mm). Prosome L/ W 1.3 –1.4, widest at end of cephalosome, and 1.8–1.9 times longer than urosome. Cephalosome somewhat triangular in dorsal view, with round integumental window on dorsal surface. Genital double-somite ( Figure 2B View Figure 2 ) L/ W 0.7 –0.9, laterally expanding into angular protuberance. Genital double-somite and subsequent two urosomites with coarsely serrate hyaline frills; teeth of mid-dorsal frill of fourth urosomite ( Figure 2B,C View Figure 2 ) larger than those of lateral and ventral frills. Anal somite ( Figure 2B,C View Figure 2 ) with row of spinules along distal margin except on dorsal and medial sides.
Caudal rami ( Figure 2B View Figure 2 ) L/ W 1.1 –1.2, distally with rows of spinules near base of dorsal terminal seta and on lateral margin; proximal dorsolateral, lateralmost terminal and medialmost terminal setae about as long as ramus; dorsodistal seta about 2.0 times longer than ramus; lateral median seta 0.9 times as long as urosome; median terminal seta 2.0 times longer than urosome. Middle terminal setae ( Figure 2D View Figure 2 ) almost homogeneously setulose, but setules on medial seta distally longer and hair-like.
Antennule ( Figure 2E View Figure 2 ) six-segmented, reaching midlength of cephalosome, with setal formula (ae indicates aesthetasc): I59, II512, III55 +spine, IV56+ae, V52 , VI510 +ae; first segment with two rows of spinules ventrally; spinules of distal row longer than those of proximal one; fourth segment longest and L/ W 1.3 .
Antenna ( Figure 2F View Figure 2 ) three-segmented, consisting of coxobasis and twosegmented endopod, with setal formula 3, 1, 12; coxobasis proximally with tuft of long hairs on anterior surface and two rows of short hairs medially; Enp2 slightly longer than Enp1, with proximal row of spinules and two distal spinules on anterior surface and intermittent row of spinules along lateral margin.
Mandible ( Figure 2G View Figure 2 ) with two unequal setae on reduced palp, and two rows of spinules near palp and base of segment, respectively.
Maxillule ( Figure 3A View Figure 3 ) consisting of praecoxal arthrite and one-segmented palp; praecoxal arthrite medially with strong spine at midlength, short seta distal to spine, and five spinules around spine, and with four strong spines apically; palp with one proximal and six apical setae.
Maxilla ( Figure 3B View Figure 3 ) incompletely four-segmented; praecoxa and coxa partly fused on posterior surface; praecoxa with two setae on endite; coxa with two setae on medial margin and on distal endite, respectively, strong spine fused to endite, and setule on spine; basis medially expanding into strong naked claw, with one-sidedly toothed strong spine and medial seta; endopod with seta, two one-sidedly toothed claw-like spines, and few spinules at socket of spine.
Maxilliped ( Figure 3C View Figure 3 ) two-segmented; syncoxa with three setae, two of them inserted at about midlength of medial margin and one at distal one-quarter on anterior surface, and row of fine spinules on distolateral corner; endopod L/ W 1.3, with five setae.
P1–P4 ( Figure 3 View Figure 3 D–G) with three-segmented rami. The spine and seta formulae as in Table 1. P1 ( Figure 3D View Figure 3 ) intercoxal sclerite with hairs along the edge of the round protuberance on each side; coxa with row of long spinules along distolateral margin on anterior side; basis with medial spine reaching beyond tip of Enp3 and long spinules near base of endopod on anterior surface; spine on Exp2 naked; proximal two spines of Exp3 naked, 1.2–1.3 times longer than segment and about twice as long as distal spine. P2–P4 intercoxal sclerite without hairs; bases with row of long spinules near base of endopod on anterior surface; proximalmost medial seta of Enp3 spiniform on distal one-third. P4 Enp3 ( Figure 3G View Figure 3 ) L/ W 1.2; medial apical spine 2.0 times longer than segment and lateral apical spine; distomedial seta spiniform as in medial apical spine but with few short setules proximally.
P5 exopod ( Figure 3H View Figure 3 ) L/W about 1.1, with two lateral and one medial spines, apical seta, oblique rows of long spinules on lateral and medial sides, and smaller spinules around sockets of spines; length ratio of three spines (from lateral to medial) to segment 1.1:1.0:1.2; seta about 1.5 times longer than segment.
Remarks
Halicyclops continentalis sp. nov. is identical to H. sinensis described by Tai and Chen (1979) in all diagnostic features. The species is distinguishable from most other congeners by having angular protuberances on the genital double-somite, the 3.4.3.3 spine formula on the P1–P4 Exp3 and characteristic spines on the P1 Exp3, of which the proximal two are much longer than the distal one. The hitherto known species and subspecies with the same formula are H. brevispinosus meridionalis Herbst, 1953 , H. b. psammophilus Pleşa, 1959, H. laminifer Herbst, 1982 , H. exiguus Kiefer, 1934 described by Defaye and Dussart (1988), H. tetracanthus Rocha, 1995 , and H. eberhardi De Laurentiis et al., 2001 . Among these, H. laminifer is the most similar to H. continentalis in having similar spines on the P1 Exp3, of which the proximal two are about twice as long as the distal one ( Herbst 1982). However, H. laminifer differs from H. continentalis by a squarely produced hyaline frill along the posterodorsal margin of the fourth urosomite and a round lateral protuberance of the genital double-somite. The other species with the 3.4.3.3 spine formula are also distinguishable by longer female antennules, of which the L/W of the fourth segment is at least 2.0 ( Herbst 1953; Pleşa 1959; Defaye and Dussart 1988; Rocha 1995; De Laurentiis et al. 2001), in contrast to 1.3–1.6 in H. continentalis , and by the absence of a triangular protuberance on the lateral surface of the genital doublesomite.
Halicyclops sinensis was described by Kiefer (1928) as a freshwater form from China that was previously described but not definitively identified by Burckhardt (1913) as ‘‘ H. aequoreus (Fischer, 1860) or a closely related new species.’’ According to Burckhardt’s description and illustration, the spine formula of the P1–P4 of the specimen is 3.4.4.3 and the medial spine on the P1 basis reaches only to the midlength of the Enp3, indicating that H. sinensis described by Tai and Chen (1979) from Chinese brackish waters differs from Kiefer’s original description ( Kiefer 1928).
Ishida (1993, 2002) also described H. sinensis from Japan. He noted that there were no morphological differences among his specimens from Hokkaido and Malaysia, and that the description in Tai and Chen (1979) provided no information on the spine number of P3 Exp3. We re-examined Ishida’s H. sinensis female specimens from Hokkaido ( Ishida 1993), which were deposited in the National Science Museum, Tokyo (NSMT Cr-11269, 11270). Results showed that this specimen had a 3.4.4.3 spine formula and that posterior margins of the urosomites lacked the coarsely serrate frills, which are present in Burckhardt’s specimen. This indicated that H. sinensis described by Ishida (1993, 2002) is neither Kiefer’s original ( Kiefer 1928) nor the present species.
The detention pond of Isahaya Bay, in which the type locality is located, was closed off by a 7-km long dike in April 1997. Ueda et al. (2003) performed plankton surveys in the pond seven times from May 1997 to July 2000 to investigate changes in plankton in relation to desalination. Mean surface water salinity in the pond steeply declined from 27 psu in May 1997 to 3.8 psu on 28 August 1997, and was almost constant at around 1 psu after April 1999. Halicyclops continentalis was first observed on 28 August 1997, when it was found densely aggregated at a single site with a salinity of 1.05 psu. At this time, copepod densities of H. continentalis were at their highest level (6.9 individuals per litre), and they continued to occur at lower densities until the end of the survey (Ueda, unpublished data). This species was also collected from the Kashima-gawa, Rokkaku-gawa, Kase-gawa, Okinohata-gawa and Yabe-gawa Rivers in the innermost area of the bay. The salinity range of these waters was 1–10 psu. There were somewhat wide variations in body length between the localities: 0.63 mm from the Rokkaku-gawa River, 0.79 mm from the Kashima-gawa River and 0.96 mm from the Yabe-gawa River. The head of the specimen from the Yabe-gawa River was rounded rather triangular.
The Chinese population of the present species, described as H. sinensis , is distributed widely from Guangzhou to Tianjin ( Tai and Chen 1979), whereas it is restricted to Ariake Bay in Japan. This geographical pattern indicates that the Ariake Bay population is probably a continental relict of the East Asian continent, as previously discussed for the brackish-water copepods Sinocalanus sinensis (Poppe, 1889) ( Hiromi and Ueda 1987) , Tortanus derjugini Smirnov, 1935 ( Ohtsuka et al. 1995) and Acartia ohtsukai Ueda and Bucklin, 2006 ( Ueda and Bucklin 2006) in this bay.

| NSMT |
National Science Museum (Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halicyclops continentalis
| Ueda, Hiroshi & Nagai, Hidefumi 2009 |
Halicyclops continentalis
| Ueda & Nagai 2009 |
H. continentalis
| Ueda & Nagai 2009 |
H. continentalis
| Ueda & Nagai 2009 |
H. continentalis
| Ueda & Nagai 2009 |
Halicyclops continentalis
| Ueda & Nagai 2009 |
H. eberhardi
| De Laurentiis 2001 |
H. tetracanthus
| Rocha 1995 |
H. laminifer
| Herbst 1982 |
H. laminifer
| Herbst 1982 |
H. laminifer
| Herbst 1982 |
Halicyclops sinensis
| , Tai and Chen 1979 |
H. sinensis
| , Tai and Chen 1979 |
Halicyclops sinensis
| , Tai and Chen 1979 |
H. sinensis
| , Tai and Chen 1979 |
H. sinensis
| , Tai and Chen 1979 |
H. sinensis
| , Tai and Chen 1979 |
H. sinensis
| , Tai and Chen 1979 |
H. brevispinosus meridionalis
| Herbst 1953 |
H. exiguus
| Kiefer 1934 |