
Parahelice pilosa ( Sakai, Türkay & Yang, 2006 ), in Sakai et al., 2006
|
publication ID |
https://doi.org/10.11646/zootaxa.4728.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:E2493FB9-5082-40A5-9408-54F3645D53C3 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB87E9-A04E-F645-FF54-FBF0257EFEEB |
|
treatment provided by |
Plazi |
|
scientific name |
Parahelice pilosa ( Sakai, Türkay & Yang, 2006 ) |
| status |
|
Parahelice pilosa ( Sakai, Türkay & Yang, 2006) View in CoL
( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 )
Pseudohelice (Parahelice) pilosa Sakai, Türkay & Yang, 2006: 51 View in CoL , figs. 61, 62, 67, 72, 84, 85 (type locality: Balayan Bay, Luzon, the Philippines) (the Philippines: Balayan Bay; Panay Island ; Indonesia: Timor; Maluku; Flores) .
Parahelice View in CoL pilosa— Ng et al. 2008: 227 (list).
Pseudohelice subquadrata — Li & Chiu 2013 View in CoL : lower image on p. 61. Not Pseudohelice subquadrata (Dana, 1851) View in CoL .
Material examined. Taiwan: 4 ♂♂ (11.3–13.8 mm) , 4 ♀♀ (15.6–18.2 mm) (NCHUZOOL 15699), Baoli R. estu- ary, Pingtung, coll. J.-H. Lee and W.-C. Wang, 1 Mar. 2012 ; 1 ♂ (16.5 mm) (NCHUZOOL 15698), Baoli R. estuary, Pingtung, 17 May 2012 ; 1 ♀ (19.7 mm) ( NMMBCD 4048 ), Baoli R. estuary, Pingtung, coll. J.-J. Li, 28 Mar. 2012 ; 1 ♂ (17.9 mm) ( NMMBCD 4357 ), Baoli R. estuary, Pingtung, coll. J.-J. Li, 8 June 2012 ; 1 ♂ (16.2 mm) ( NCHU- ZOOL 15718 ), Baoli R. estuary, Pingtung, coll. J.-J. Li, 5 Aug. 2012 ; 1 ♂ (16.0 mm), 1 ♀ (14.2 mm) (NCHUZOOL 15693), Gangkou R. estuary, Pingtung, coll. J.-H. Lee and W.-C. Wang, 29 Jan. 2012 ; 1 ♂ (16.4 mm) (NCHUZOOL 15696), Gangkou R. estuary, Pingtung, 10 May 2012 ; 2 ♂♂ (14.5–16.3 mm), 6 ♀♀ (15.3-17.3 mm) (NCHUZOOL 15700), Gangkou R. estuary, Pingtung, coll. J.-H. Lee and W.-C. Wang, 30 Mar. 2012 ; 1 ♂ (17.9 mm) (NCHUZOOL 15719), Gangkou R. estuary, Pingtung, coll. J.-H. Lee, 21 June 2012 ; 2 ♂♂ (11.9–19.0 mm) (NCHUZOOL 15689), Gangkou R. estuary, Pingtung, coll. J.-J. Li, 7 July 2012 ; 1 ♂ (16.1 mm) (NCHUZOOL 15691), Gangkou R. estu- ary, Pingtung, 11 July 2012 ; 1 ♂ (15.0 mm) (NCHUZOOL 15732), 1 ♀ (17.7 mm) (NCHUZOOL 15729), Gangkou R. estuary, Pingtung, coll. J.-H. Lee, 21 Nov. 2012 . Indonesia: 1 ♀ (7.0 mm) (NCHUZOOL 15721), Tanah Lot, Beraban , Bali, 22 July 2014 .
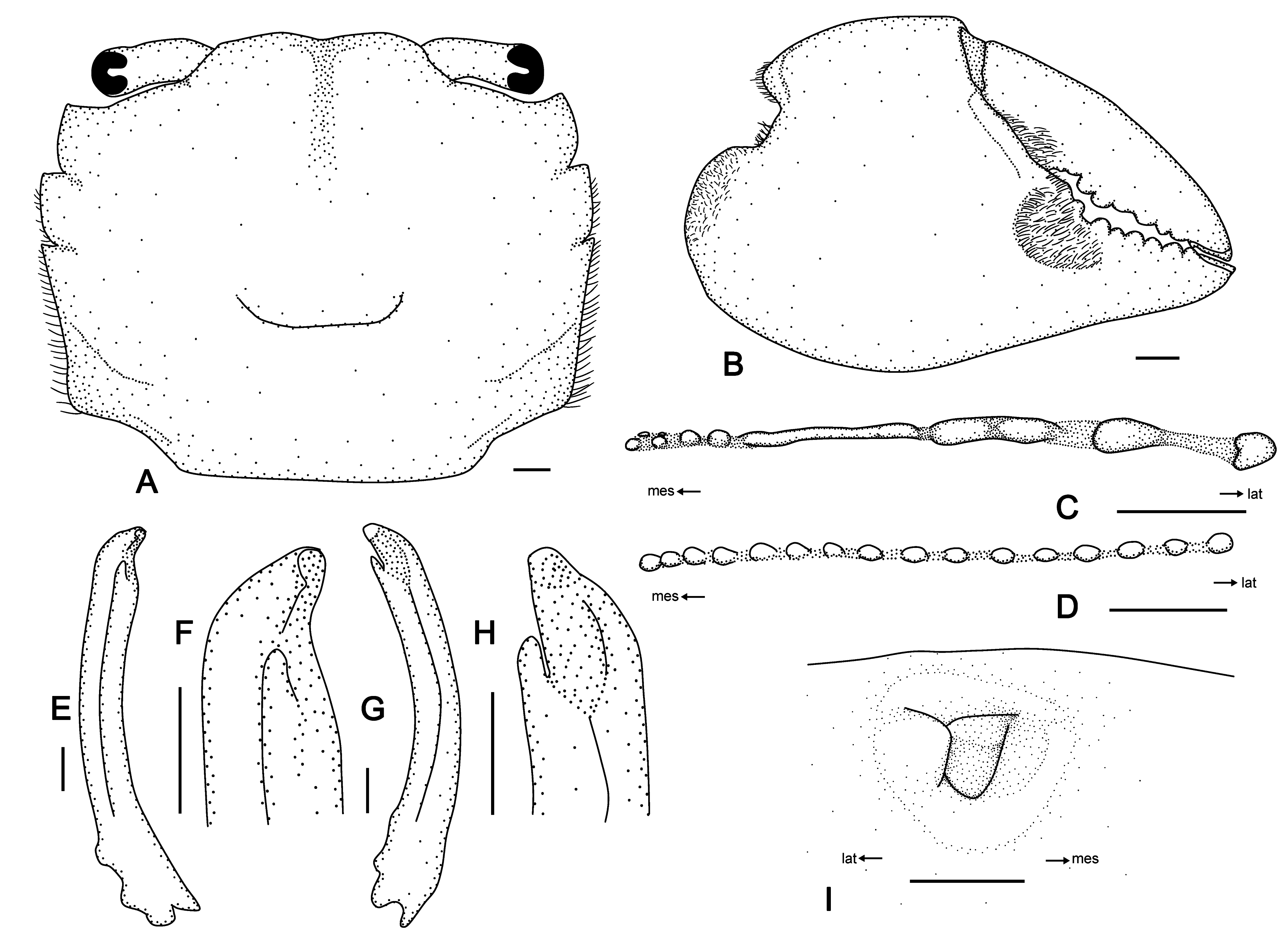
Diagnosis. Carapace quadrate, slightly broader than long, 1.19-times as broad as long (n = 48); surface convex, weakly punctate, granulate, with distinct groove between epigastric regions. Frontal margin slightly concave. Anterolateral margin with 3 teeth including orbital tooth; last tooth weak, indistinct. Infraorbital ridge in male heteromorphic, proximal part with 4–7 rounded tubercles, followed by elongated, laterally slightly inflated crest, distal part with 2 rounded tubercles; female with 14–22 concentrated isomorphically interspaced, rounded tubercles. Chelipedal palm stout, surface almost smooth; usually unequal in male, distinct patch of setae at base of fingers, mostly expanding onto fixed finger; chelipeds in female usually equal, glabrous. Ambulatory legs broad, anterior margins of merus, carpus, and propodus covered with short setae. Male G1 broad, stout, slightly curved towards distal end; female vulvae sunken in lateral part, with short semicircular operculum.
Ecological notes. Habitats include muddy substrate, with or without vegetation in southern Taiwan (at a distance of 600–800 m from a river mouth) ( Fig. 7A, B View FIGURE 7 ); and sandy mud with some grasses in Bali, Indonesia near a river mouth ( Fig. 7D View FIGURE 7 ). This species is sympatric with Par. daviei , Par. pilimana , and Pse. subquadrata in southern Taiwan, with burrow depths of ca. 50 cm or less; sympatric with Pse. subquadrata in Bali, Indonesia.
Distribution. From Taiwan (Pingtung), the Philippines (Luzon, Panay) to Indonesia (Timor; Maluku; Flores; Bali). This species is a new record for Taiwan and Bali.
Remarks. Although we did not examine specimens of Par. pilosa from its type locality (Balayan Bay, Luzon I.) or the adjacent area, our specimens from Taiwan and Indonesia agree well with the original description ( Sakai et al. 2006), including carapace ( Fig. 6A View FIGURE 6 ; Sakai et al. 2006: figs. 84, 85), male chelae ( Fig. 6B View FIGURE 6 ; Sakai et al. 2006: fig. 84), male infraorbital ridges ( Fig. 6C View FIGURE 6 ; Sakai et al. 2006: fig. 61), and G1s ( Fig. 6 View FIGURE 6 E–H; Sakai et al. 2006: fig. 67).
The morphology of female Par. pilosa is similar to that of female Pse. subquadrata in the mature size, carapace, chelipeds, and ambulatory legs; there is also no patch of setae at the base of the female fingers. Both species are sometimes sympatric in Taiwan, and it is quite difficult to identify females. Based on the female specimens examined, a useful character for distinguishing females is the infraorbital ridge, which has 14–22 concentrated isomorphically interspaced and rounded tubercles in female Par. pilos a ( Fig. 6D View FIGURE 6 ), but is composed of a striated crest proximally and two or three large tubercles distally in Pse. subquadrata ( Sakai et al. 2006: fig. 58).
In life, Par. pilosa always has a distinct “bi-colored” carapace in adults, i.e., dark-brown in the anterior region and light-orange in the posterior region ( Fig. 5E View FIGURE 5 ). Following this character, one of the figures of “ Pse. subquadrata ” in Li & Chiu (2013: lower image on p. 61) is probably Par. pilosa instead.
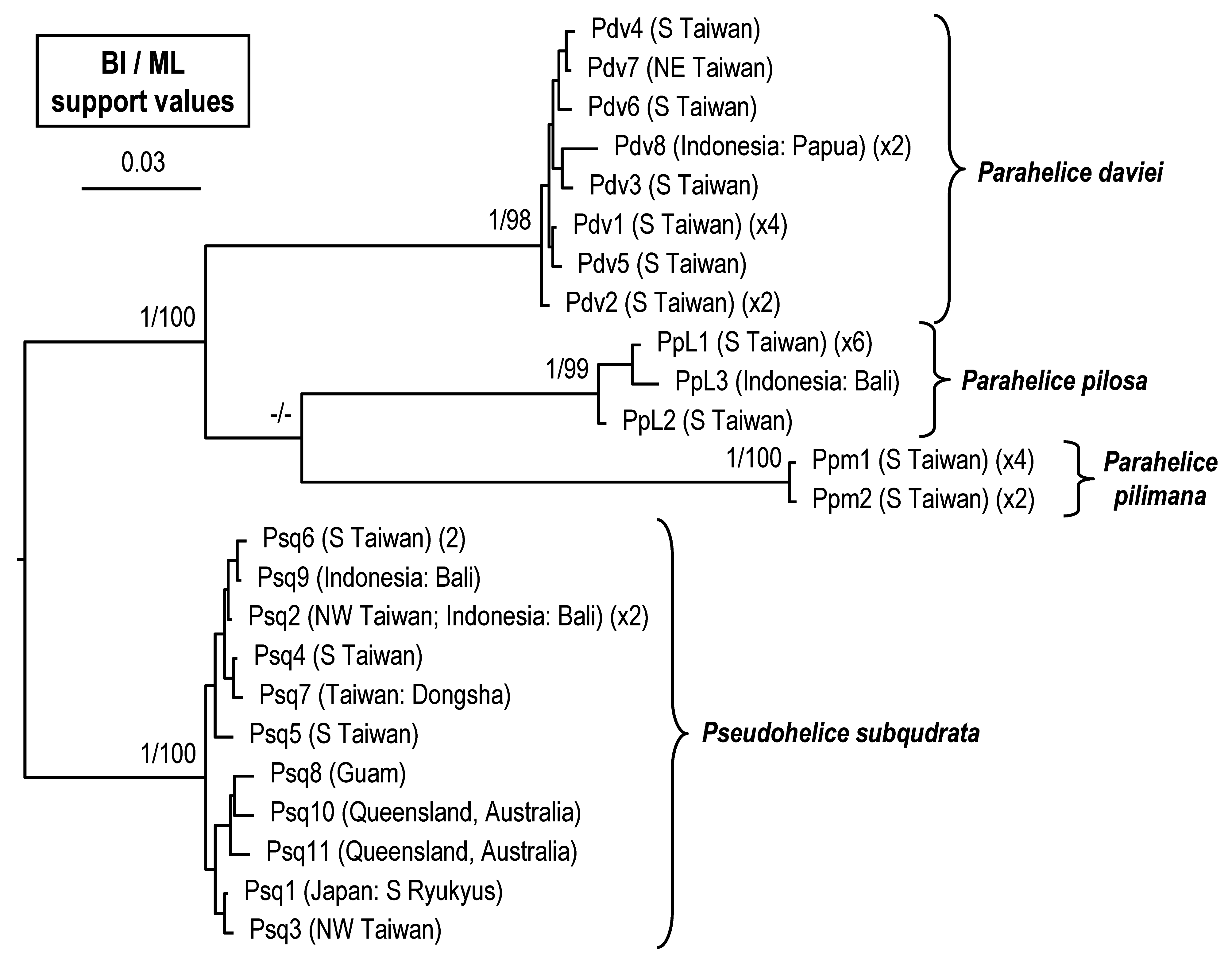
Molecular analyses. Molecular results of the COI marker comprised 27 specimens of three species of Parahelice , as well as 12 specimens of Pse. subquadrata ( Table 1 View TABLE 1 ). The reconstructed phylogenetic tree, with support values from the BI and ML analyses ( Fig. 8 View FIGURE 8 ), shows four well-supported clades, which corresponds to the four species in this study. The three species of Parahelice form a main clade, but the relationship among them is unresolved.
The mean pairwise nucleotide divergence of K2P distances and bp differences of haplotypes of the four species are shown in Table 3 View TABLE 3 . The intraspecific nucleotide divergences of the four species were all <1.54%, with Par. pilimana possessing the lowest intraspecific divergence (≤0.15%). The interspecific divergences among the four species were all ≥13.13%. Thus, the interspecific divergence was at least 8.5-fold greater than intraspecific values (between Par. daviei and Par. pilosa ), supporting the four species being distinct species.


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |
Parahelice pilosa ( Sakai, Türkay & Yang, 2006 )
| Shih, Hsi-Te, Hsu, Jhih-Wei, Li, Jheng-Jhang, Ng, Ngan Kee & Lee, Jung-Hsiang 2020 |
Parahelice
| Ng, P. K. L. & Guinot, D. & Davie, P. J. F. 2008: 227 |
Pseudohelice (Parahelice) pilosa Sakai, Türkay & Yang, 2006: 51
| Sakai, K. & Turkay, M. & Yang, S. - L. 2006: 51 |