
Neomysis integer ( Leach, 1814 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4142.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FA423164-276C-44B0-A417-8E97AC3DF0AA |
|
DOI |
https://doi.org/10.5281/zenodo.6088684 |
|
persistent identifier |
https://treatment.plazi.org/id/03B9BA4E-1823-FFF2-CAF6-F9B8FED54047 |
|
treatment provided by |
Plazi |
|
scientific name |
Neomysis integer ( Leach, 1814 ) |
| status |
|
Neomysis integer ( Leach, 1814) View in CoL
Fig. 20 View FIGURE 20
Short selection from 22 synonymy statements with a total of 622 references:
Praunus integer Leach, 1814 : Stebbing, 1893.
Mysis integer: Leach 1815, 1817 ; White 1847, 1850; Bell 1853 (with query).
Mysis View in CoL entier [non-Linnean assignment]: Desmarest 1823, 1825; H. Milne Edwards 1837 (in remarks).
Praunus flexuosus: Desmarest 1825 View in CoL (in synonymy).
Mysis vulgaris J. V. Thompson, 1828 : H. Milne Edwards 1837; Kröyer 1861; Czerniavsky 1882a (partim: 2, 7); Ekman 1935; Muus 1967 (partim: Tab. 29).
Mysis scoticus J. V. Thompson, 1828 .
Mysis View in CoL commun [non-Linnean assignment]: H. Milne Edwards 1837.
Neomysis vulgaris: Czerniavsky 1882a (partim: 15), 1882b, 1887; Needham 1937; Kinne 1955; Ten 1991; Mitina & Kharina 2011.
Neomysis integer: W. M. Tattersall 1912 View in CoL ; Băcescu 1941; Brun 1967; Wittmann & Ariani 2009; Remerie et al. 2004; Wittmann 2013; Zettler 2015.
Material examined (hand net, leg. K. J. Wittmann, if not stated otherwise):
Mediterranean drainage. 3 F ad. 11.6 mm (one dissected), 1 M ad. 9.1 mm, 1 M subad., 1 F imm., Canal d'Arles à Fos within the estuary of the Grand Rhône, S = 12.3, NHMW reg. no. 22995, for additional data see Wittmann & Ariani (2009); 1 M ad. 7.6 mm (dissected and mounted on slides), Canal du Rhône à Sète within the estuary of the Petit Rhône, 43.6432N 004.4298E, altitude 1 m, sea distance 30 km along canals or 42 km along the river Petit Rhône, 0.2–1.8 m depth, from shore macrophytes and from boulders with filiform algae, among masses of Atyaephyra desmarestii , v = 0 m/s, S = 0.4, 1010 µS/cm, 24.5°C, pH 7.92, 6.54 mg O2/l, 53 NTU, 19 June 2009.
NE-Atlantic drainage. 1 F ad. 8.7 mm, 8 M ad. 7.6–8.7 mm, 4 F subad., 5 M subad., 15 imm., 2 juv., France, estuary of Gironde River at Cadou, river-km K.60, 45.3097N 000.7815W, altitude 0 m, sea distance 37 km, 0.5–3 m depth, from stone walls with filiform algae, v = 0.2–0.4 m /s, S = 4.1, 7620 µS/cm, 23.6°C, pH 7.67, 6.34 mg O2/ l, 222 NTU, 22 June 2009, NHMW 25712 View Materials GoogleMaps ; 23 F ad. 8.7–11.6 mm, 31 M ad. 6.7–9.3 mm, 15 F subad., 15 M subad., 24 imm., 6 juv., France, Bay of Biscay , Bassin d'Arcachon, chenal de La Hume, 44.5921N 001.1237W, S = 1.2, 2 Aug. 1983, don. J. C. Sorbe (Arcachon), RNMH GoogleMaps . CRUS.E.3782; 1 juv., The Netherlands, Oude Rijn , Katwijk , at Valkenburgseweg , 52.1877N 004.4274E, altitude 0 m, sea distance 4 km, 0.2–1 m depth, among Nuphar , S = 0.4– 1.0, 23 July 1998; 60 F ad. 9.2–11.8 mm, 5 M ad. 9.1–10.8 mm, 2 F subad., 1 M imm., The Netherlands, Den Helder, Noordhollands kanaal, 52.9638N 004.7667E, altitude 2 m, sea distance 5 km, 1–2 m depth, among macrophytes, S = 2–3, 22 June 1989, NHMW 25450 View Materials GoogleMaps ; 26 F ad. 10.7–16.4 mm, 40 M ad. 9.0–12.0 mm, 2 F subad., 3 M subad., 49 imm., 23 juv., Scotland, Argyll , Loch Creran , near Inver , 56.5137N 005.3728W, altitude 0 m, sea distance about 26 km, 0.2–0.3 m depth, among algae, low tide, tidal current v = 0.3–0.6 m /s, S = 0–25, 11 Sept. 1983; 1 F ad., 1 M ad., 1 imm., Scotland, Argyll , Firth of Lorn, Dunstaffnage Bay, 56.450N 005.433W, 0.1–0.2 m depth, among algae and on mud, S = 28–33 (mixoeuhaline), 12°C, 13 Sept. 1983, NHMW 25449 View Materials GoogleMaps .
Baltic drainage. 16 F ad. 12.0– 16.7 mm, 61 M ad. 7.6–15.7 mm, 13 subad., 5 imm., 3 juv., Germany, Fehmarn- Belt, Puttgarden, 54.505N 011.222E, 0.3 m depth, sand, S = 14, 23 Aug. 1985, leg. K. J. Wittmann & A. P. Ariani, RNMH GoogleMaps .CRUS. E.3783; 172 F ad. 10.7–16.7 mm, 35 M ad. 8.5–12.9 mm, 10 F subad., 10 imm., Sweden, Askö Island , 58.8288N 017.6366E, 0.3–1 m depth, among algae, S = 7, 22 Aug. 1985, NHMW 25713 View Materials GoogleMaps .
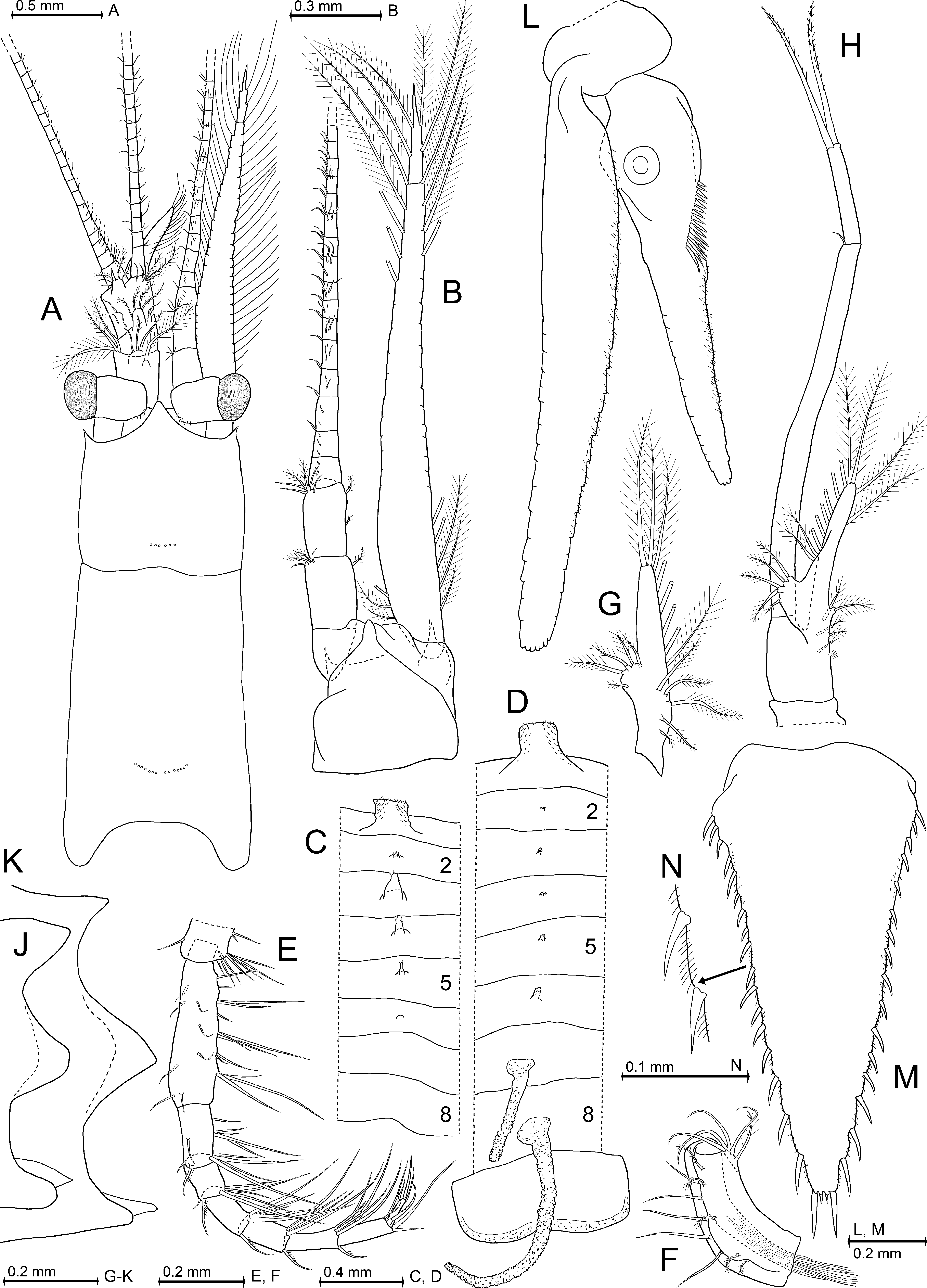
Supplementary description. Adult body length 6–19 mm, eyes normal. Antennal scale strongly elongate, setose all around, terminally pointed in both sexes ( Fig. 20 View FIGURE 20 A, B). Its two segments separated by a transverse, not always distinct suture; apical segment is 12–18% scale length. Carapace ( Fig. 20 View FIGURE 20 A) ends anteriorly in an apically acute, triangular rostrum; no subrostral plates; lateral margins of carapace produced into a pair of anteriorly directed spiniform processes; dorsal face of carapace with 5–12 cervical pores in about linear, transversal arrangement; and with 12–29 cardial pores showing a transverse arrangement in two about symmetrical, slightly curved subgroups. Thoracic endopod 8 represents the strongest leg in both sexes by being always more stout and by being 0–30% longer compared with the subequal endopods 3–7; carpopropodus of endopods 3–8 with 5–6, 5–7, 5–7, 5–7, 5–7, or 6–8 segments, respectively (adult males tend to show more segments than same-sized adult females); these endopods ending in short, weak, slightly bent claws ( Fig. 20 View FIGURE 20 E). Flagellum of thoracic exopods 1 and 8 each with 8 segments, that of exopods 2–7 each with 9 (10) segments, not counting the large intersegmental joint between the 1-segmented basis and its multi-segmented flagellum. Outer distal corner of basis always ending in a small spine.
First thoracic sternite produced into an anteriorly directed, distally hairy (with minute setae), terminally nearly quadrangular, medial lobe in both sexes ( Fig. 20 View FIGURE 20 C, D). Adult males ( Fig. 20 View FIGURE 20 C): thoracic sternites 2–5 each with small median, terminally blunt processes each bearing 2–4 minute acute scales; sternite 6 medially with transverse cuticular ridge that may represent a vestigial process; sternites 7, 8 with smooth median portions. Adult females ( Fig. 20 View FIGURE 20 D): thoracic sternites 2–6 medially with vestigial processes; a total of two large, finger-like processes project from thoracic sternites 7, 8 ventrally into the marsupium; these processes show densely set knob-like structures giving their surface a moruloid appearance, no scales, large longitudinal tube inside each process. Marsupium supported by a large, backwardly produced plate ( Fig. 20 View FIGURE 20 D) from last thoracic sternite. This plate present only in females and represents an apron which overlaps part of first pleon sternite. Its alignment with anterior margin of pleon suggests a function as mechanical protection of marsupium upon flipping of pleon, which occurs most violently upon sudden escape. All pleopods reduced, fused and styliform ( Fig. 20 View FIGURE 20 G) in both sexes, with exception of male pleopod 4 ( Fig. 20 View FIGURE 20 H). This latter pleopod biramous, strongly elongate; its 2-segmented exopod reaches beyond base of uropods. Scutellum paracaudale mostly subtriangular with convex upper and concave lower margins ( Fig. 20 View FIGURE 20 K), less frequently biconvex, and even less frequently sinusoid ( Fig. 20 View FIGURE 20 J); tip blunt to well rounded. Endopods of uropods with 14–55 spines arranged in dense series along proximal half of inner margin ( Fig. 20 View FIGURE 20 L). Statoliths composed of fluorite (according to Lowenstam & McConnell 1968; confirmed by own determinations on material from Canal d'Arles à Fos, chenal de La Hume, and Askö Island). Telson ( Fig. 20 View FIGURE 20 M) entire, subtriangular, longer than ultimate pleonite; lateral margins all along with continuous series of spines, numerous fine hairs between the lateral spines ( Fig. 20 View FIGURE 20 N); narrow, truncate apex with a pair of short median spines flanked by a pair of long latero-apical spines.
Distribution ( Fig. 16 View FIGURE 16 ). Common in shallow coastal waters of the temperate to boreal NE-Atlantic from southern Spain to Norway, also common in the Baltic. Also found in the White Sea and the Barents Sea ( Zimmer 1933, Holmquist 1972, Petryashov 2004). A presumably disjunctive population was detected by Băcescu (1941) in the NW-Mediterranean (Rhône Delta). This species occurs from anhaline to hyperhaline conditions, yet mainly in oligo- to polyhaline waters, where it has strong osmoregulatory capabilities (at S = 3–32 according to Vilas et al. (2006)). Freshwater records were made in waters pertaining to the drainage basins of the NE-Atlantic ( Scott 1894, Elton 1937, Tattersall & Tattersall 1951, Hynes et al. 1960, Ketelaars et al. 1999, Lee & Bell 1999, Braune 2004) and of the Baltic ( Gasiunas 1965, 1972; Zettler 1998).
NE-Atlantic populations of N. integer are commonly found from oligohaline to polyhaline conditions, with main occurrence in the mesohaline reach, rarely in fresh-water. One own sample (listed above) of a juvenile was taken in almost fresh-water (S = 0.4–1.0) in an old Rhine branch at the coast of The Netherlands. The abundance of this species along the Westerschelde estuary at the Dutch-Belgian border is primarily correlated with salinity; here it lives at S = 0.6–27 throughout the year ( Rappé et al. 2011). Similarly, Roast et al. (1998) observed a minimum of S = 1 for this species in a Cornwall estuary. There is a number of additional near-freshwater records ( Castel 1993, Bernát et al. 1994, Köpcke & Kausch 1996, David et al. 2005, Vilas et al. 2009) as well as the above-cited true freshwater records along the Atlantic coasts of Europe.
As a first freshwater record for the Mediterranean, we documented one adult male at S = 0.4 in a canal pertaining to the estuary of the Petit Rhône ( Fig. 16 View FIGURE 16 ; see material list above). Brun (1967) reported this species from the chlorinity range of 0.5–5‰ (salinity 0.9–9) in the estuary of the Grand Rhône. Wittmann & Ariani (2009) found it at S = 12.3 in the Canal d'Arles à Fos at a station within this estuary. Aguesse & Bigot (1960) reported N. integer together with Mesopodopsis slabberi from the Étang Baisse Salée (Camargue, Rhône delta). The salinities of this water body ranged from S = 0.8–40.1 (mostly 3–8) in 1955–1958. However, the exact range experienced by the mysids was not specifically indicated. For potential pathways of areal expansion to the Mediterranean, see ‘Discussion’.

| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Mysinae |
|
Tribe |
Mysini |
|
Genus |
Neomysis integer ( Leach, 1814 )
| Wittmann, Karl J., Ariani, Antonio P. & Daneliya, Mikhail 2016 |
Neomysis integer:
| W. M. Tattersall 1912 |
Neomysis vulgaris:
| Czerniavsky 1882 |
Mysis vulgaris
| J. V. Thompson 1828 |
Mysis scoticus
| J. V. Thompson 1828 |
Praunus flexuosus:
| Desmarest 1825 |
Mysis integer:
| Leach 1815 |
Praunus integer
| Leach 1814 |