
Oligobregma bakkeni, Mendes & Paiva & Rizzo, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5424.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:3DF7F8AA-BA4C-4C78-BB1E-450537B79CB6 |
|
DOI |
https://doi.org/10.5281/zenodo.10815160 |
|
persistent identifier |
https://treatment.plazi.org/id/03B887F7-3E09-6F7A-F884-B22E00D4FBFE |
|
treatment provided by |
Plazi |
|
scientific name |
Oligobregma bakkeni |
| status |
sp. nov. |
Oligobregma bakkeni sp. nov.
https://zoobank.org/NomenclaturalActs/ F5A5522E-AA2B-4B26-B238-F9FC339A3CAB
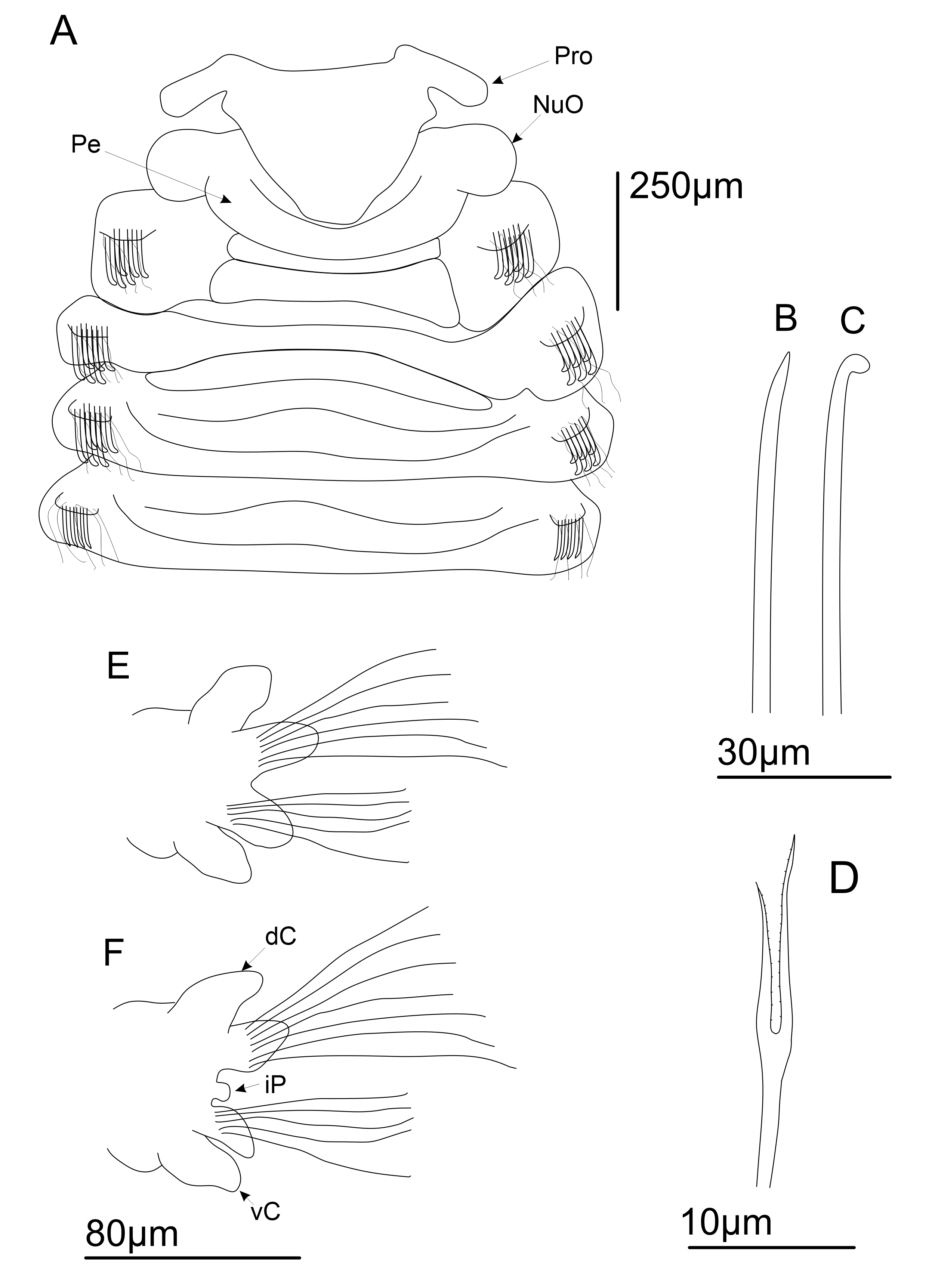
Figures 6 View FIGURE 6 & 7 View FIGURE 7
Type material. UERJ9225 (Holotype: Ambes5; A9 R1 ; Lat: -21.161194; Long: -38.873622; 04 Jan 2012; 2498 meters deep) . UERJ9226 (Paratype: Ambes5; D9 R1 ; Lat: -20.576756; Long: -38.681556; 06 Jan 2012; 2460 meters deep) .
Description. Holotype adult but incomplete, lacking posterior end. Measuring 8 mm long, 2.5 mm wide on expanded region and 1 mm wide on narrowest region, for 18 chaetigers. Moderate sized species, Paratype measuring 10 mm long, 1 mm wide on expanded region and 0.5 wide on narrowest region, for 23 chaetigerous segments. Both holotype and paratype incomplete, lacking posterior end. Body arenicoliform, weakly expanded over chaetiger 3–8. Colour in alcohol yellowish. Body surface covered by secondarily annulated rings. Secondary annuli smooth in both anterior and posterior regions.
Triangular prostomium, with two long horns, both projected laterally ( Figs 6A View FIGURE 6 ; 7A View FIGURE 7 ). Eyes absent. Nuchal organs observed, present as dorso-lateral saccular projections between prostomium and peristomium. Peristomium achaetous, uniannulated dorsally and ventrally. Proboscis smooth ( Fig. 7B View FIGURE 7 ). Ventral groove present from chaetiger 2, with quadrangular uni- to triannulated pads from chaetiger 2, forming a ventral mid-ridge up to end of body. Each pad paired to a single chaetiger. Posterior pads smaller and narrower than anterior ones. Dorsally and ventrally, chaetiger 1 presents two secondary annuli; from chaetiger 2, three annuli connect to parapodial lobe, plus one superior and inferior annulation delimitating segment.
Dorsal and ventral cirri present from chaetigers 13–14 ( Figs 6E–F View FIGURE 6 ; 7C View FIGURE 7 ). Interramal papillae present ( Fig. 7C View FIGURE 7 ); rounded when everted, observed from chaetiger 13, filled with granular content. Dorsal and ventral cirri present as a broad basis transitioning smoothly to a rounded tip ( Figs 6E–F View FIGURE 6 ; 7C View FIGURE 7 ). Strong and numerous blackish tubular glands may be present in all cirriferous chaetigers; sparse and less numerous initially, then densely entangled, filling all internal space posteriorly ( Fig. 7C View FIGURE 7 ).
Parapodial lobes reduced in anterior chaetigers ( Fig. 7B View FIGURE 7 ), becoming evident from midbody to posterior chaetigers ( Figs 6E–F View FIGURE 6 ; 7C View FIGURE 7 ). Notopodial and neuropodial lobes differing in size. Notopodial lobe always larger and longer than neuropodial lobe ( Figs 6E–F View FIGURE 6 ; 7C View FIGURE 7 ). Both podial lobes of same shape on midbody to posterior chaetigers. Parapodial lobes initially present as inconspicuous asymmetrical elevations on anterior to midbody chaetigers; then become longer and wider, emerging as a broad elevation transitioning to an apically rounded tip, from first cirriferous chaetigers on midbody ( Fig. 6E View FIGURE 6 ). On posterior chaetigers, lobes becoming longer and thinner, emerging as a broad basis abruptly transitioning to a pointed tip, forming a “knife-like” asymmetrical structure ( Figs 6F View FIGURE 6 ; 7C View FIGURE 7 ).
Acicular spines present from chaetiger 1–4 on noto and neuropodia ( Figs 6B–C View FIGURE 6 ; 7D–E View FIGURE 7 ). On notopodia, chaetigers 1–2 present two anterior rows of strong acicular spines, with a terminal curved tip, plus a posterior row of capillary chaetae ( Figs 6C View FIGURE 6 ; 7D View FIGURE 7 ); numbering 9–10 spines on anterior and posterior rows. Chaetiger 3 presents on noto and neuropodia a single anterior row of 9–10 acicular spines, with the same shape of chaetigers 1–2, plus two posterior rows of capillaries. Chaetiger 4 presents one anterior row of nine straight and apically pointed acicular spines, plus two posterior rows of capillaries ( Figs 6B View FIGURE 6 ; 7E View FIGURE 7 ).
Short spinous chaetae absent. Lyrate chaetae present from chaetiger 3 ( Figs 6D View FIGURE 6 ; 7F View FIGURE 7 ), numbering 4 in each fascicle on anterior chaetigers, reaching 8–11 in number on midbody chaetigers, then 4–5 on posterior chaetigers; always with subequal tynes (tyne’s ratio = 1.2) and in a single row anterior to capillaries. Anterior lyrate chaetae smaller, becoming longer on midbody to posterior chaetigers. Capillaries organized in one row on chaetigers 1–2, then two rows from chaetiger 3 up to posterior chaetigers. Pygidium nature unknown.
Remarks. Although belonging to the subgroup of Oligobregma species presenting acicular spines from chaetigers 1–4 in both rami, O. bakkeni sp. nov. differs from its congeners by the presence of acicular spines terminating with rounded curved tips on chaetigers 1–3, changing to transitional acicular spines on chaetiger 4. Only two types of acicular spines are reported within Oligobregma : curved or hirsute, both with or without terminal arista on their apical ending.
Finally, the prostomium and its horns are also unique in this species, differing from all its congeners on the trapezoidal shape, “shield-like”, accompanied by long, and thin horns, these reaching the half of prostomium in length, always laterally directed. In O. bakkeni sp. nov., lyrate chaetae co-occur with spinous chaetae, emerging from chaetiger 3. None of the species with acicular spines on chaetiger 1–4 has lyrate chaetae from chaetiger 3. However, species with acicular spines on chaetiger 1–2 possess lyrate chaetae emerging from chaetiger 3, but never co-occurring with acicular spines as in O. bakkeni sp. nov.
Distribution. O. bakkeni sp. nov. was found living on Espírito Santo basin continental slope, from a bathymetric range of 2460–2498 meters deep, predominantly among muddy sediment.
Etymology. The specific epithet “ bakkeni ” was chosen to honor to Dr. Torkild Bakken, for his important contributions to Scalibregmatidae taxonomy and annelids in general.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.