
Marmosops Matschie, 1916
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2001)263<0003:TMOPFG>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03B69D69-FFE6-376C-8508-FE6EFBD2FA32 |
|
treatment provided by |
Marcus |
|
scientific name |
Marmosops Matschie |
| status |
|
Marmosops Matschie View in CoL
Thirtyfive voucher specimens from Paracou are referable to Marmosops parvidens in the sense of Pine (1981), but close examination of pelage characters and correlated morphometric variation in our sample indicates that two species are present. The following paragraphs summarize the evidence supporting this conclusion.
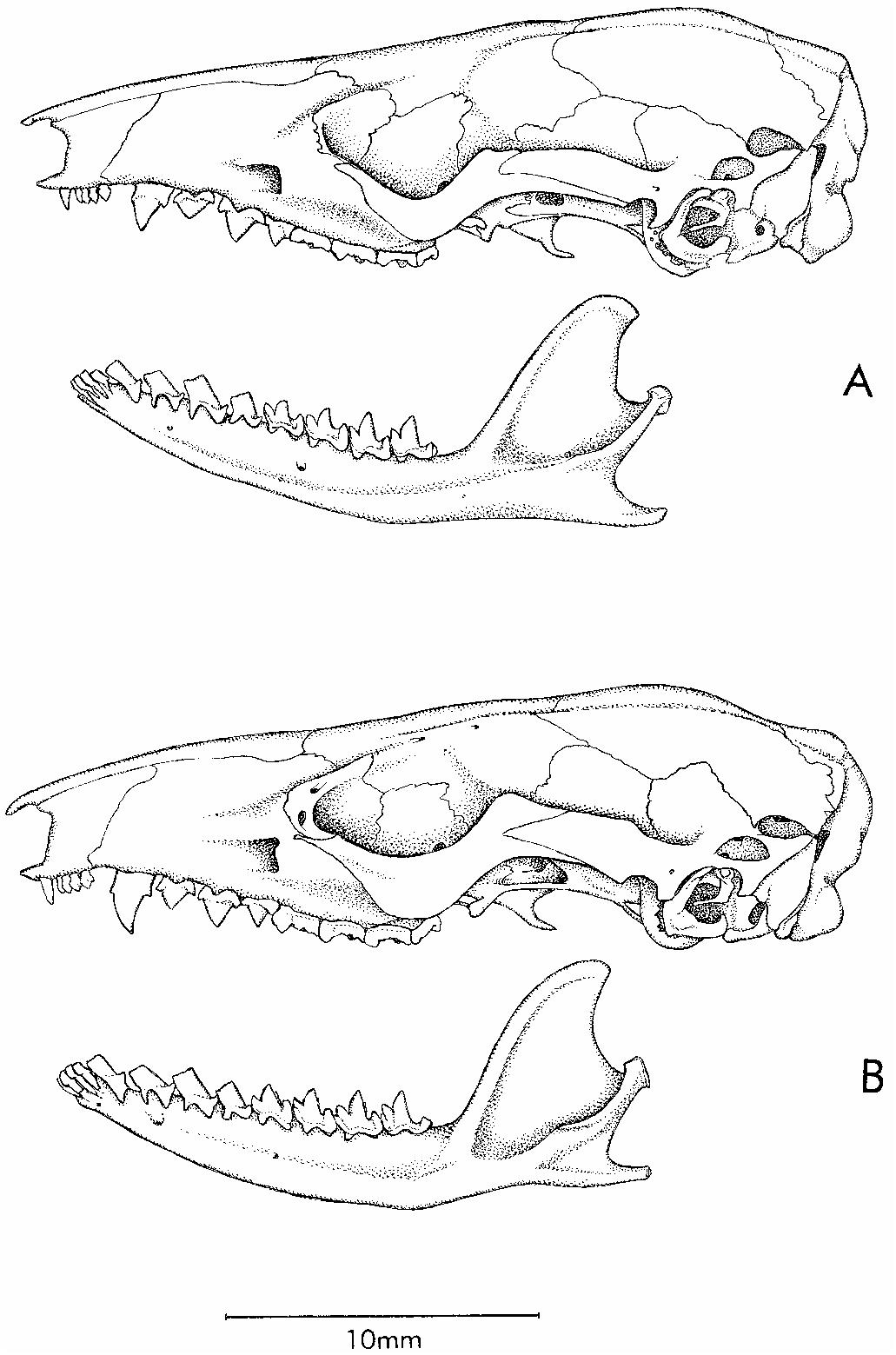
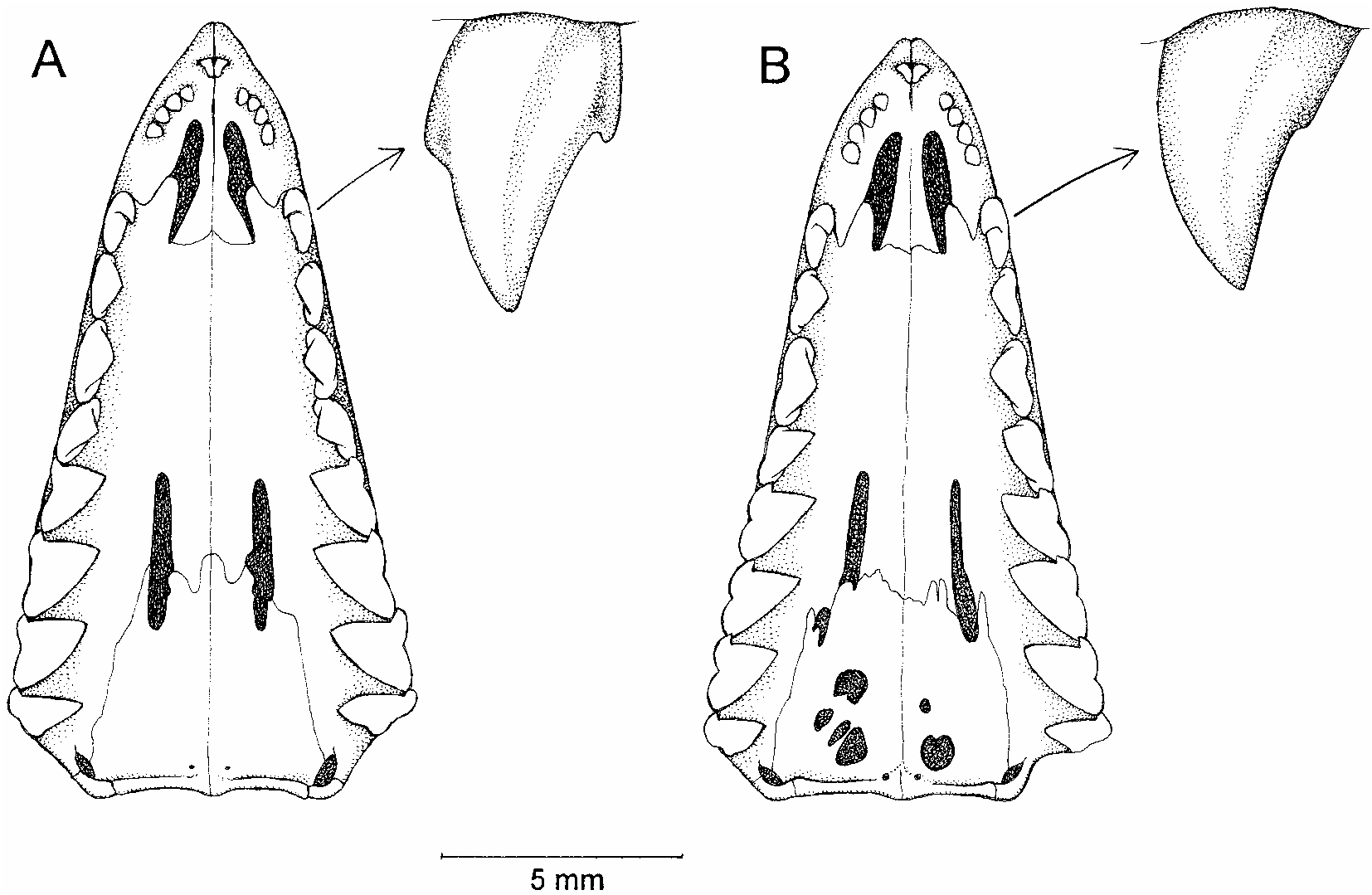
All of the specimens in question are small opossums (21–33 g adult weight) with dark facial masks, dull reddishbrown or grayishbrown dorsal fur, small fore and hindfeet (each with diminutive claws and six separate plantar tubercles), and long tails. The scrotal sacs of males are entirely white. Female specimens lack any trace of a pouch and appear to have have 3–1–3 = 7 or 4–1–4 = 9 inguinalabdominal mammae (fig. 15B). Oth er distinctive attributes include: the grossly enlarged central hair of each caudalscale triplet; a prominent, spoonshaped, fleshy tubercle supported internally by bone on the lateral aspect of the wrist of males (fig. 20); smoothly rounded supraorbital margins without distinct beads or processes (fig. 19A); conical alisphenoid bullae with prominent anteromedial processes (fig. 21); absence of posteromedial (palatine) palatal vacuities (fig. 22); upper canines with distinct anterior and posterior accessory cusps; and lower canines that resemble the lower premolars in shape and size, forming a moreorless undifferentiated series of four subequal teeth (fig. 23). Some of these traits are common to many ‘‘marmosines’’, others are perhaps diagnostic of the genus Marmosops , and a few may define a distinctive group of species closely related to M. parvidens . For present purposes, however, these attributes serve to distinguish examples of parvidens like Mar mosops from all of the other marsupial taxa that we collected at Paracou.
Within this series, the most conspicuous external variation involves fur color. We scored three pelage characters for statistical analysis. Dorsal coloration was classified as ‘‘fuscous’’ (dusky grayishbrown) or ‘‘reddish’’ (a subtly warmer tone) according to the predominant hue of the unruffled fur. Although this chromatic contrast disappears in material stored for years in alcohol, dried skins and fresh fluid specimens (recently removed from formalin) were readily assigned to one or the other of these two states. 7 Ven
7
At the time of writing ( 1997), specimen assignments to dorsal and ventral color classes (recorded in 1993) are difficult to verify in fluidpreserved material (the bulk of our vouchers) due to fading. Five dried skins, how
tral coloration was classified as ‘‘white’’ if the selfcolored fur and the tips of graybased hairs lacked any pigmentation, or ‘‘cream’’ if the selfcolored fur and graybased hair tips were pale yellowish. Ventral pattern was classed as ‘‘narrow’’ if selfcolored (pure white or cream) fur was confined to the midline (sometimes as a discontinuous streak) by a broad zone of graybased fur on each side, ‘‘broad’’ if almost the entire ventrum was selfcolored, or ‘‘intermediate’’ for speci
ever, retain their original colors: AMNH 266423, 267007, and 267352 have ‘‘fuscous’’ dorsal fur and ‘‘white’’ underparts; AMNH 266426 has ‘‘reddish’’ dorsal fur and ‘‘cream’’ underparts; and AMNH 267817 has ‘‘reddish’’ dorsal fur and ‘‘white’’ underparts. The extent of graybased ventral fur (another pelage character we scored for statistical analysis) is not affected by ageing or preservatives.
mens with substantial amounts of both selfcolored and graybased ventral fur.
Chisquared tests of independence provide no evidence for age or sex effects on the expression of these pelage traits. In fact, the largest agesex class in our sample (16 adult males) includes individuals with fuscous and reddish dorsal fur, specimens with white and creamcolored ventral fur, and examples of all three conditions of ventral pattern. However, pelage characters are not independently distributed inter se (table 7). Most animals with fuscous dorsal fur have selfcolored ventral fur narrowly confined to the midline or bordered by extensive lateral zones of graybased fur; by contrast, many animals with reddish dorsal fur have almost entirely selfcolored ventral fur, and none has selfcolored fur narrowly confined to the midline. Similarly, almost all animals with fuscous dorsal fur have white (or whitetipped) ventral fur, whereas about onethird of the specimens with reddish dorsal fur have cream (or creamtipped) underparts.
We used oneway ANOVAs to test for morphometric divergence between individu
TABLE 7 ChiSquare Tests for Independence of Pelage Charactersa among Marmosops Specimens from Paracou
als with red and fuscous dorsal fur among adult males (the only agesex class in our sample large enough for such analyses) and found highly significant differences (p <0.01) in length of molars, palatal breadth, zygomatic breadth, height of the upper canine (measured from the posterior accessory cusp to the tip of unworn teeth), and nasal breadth. Other external and craniodental measurements showed no significant divergence. Not surprisingly, principal components analysis of the logtransformed craniodental measurement data provides clear separation of dorsal fur color classes aligned with the first eigenvector (fig. 24), the coefficients of which (table 8) reflect essentially the same differences as those indicated by univariate statistics: fuscous adult males have taller canines, broader nasals, and wider zygomas than do reddish adult males, with smaller (but still substantial) differences in molar size and palatal breadth. Similar morphometric contrasts between dorsal fur color classes can be seen among the few adult females in our sample, and among immatures.
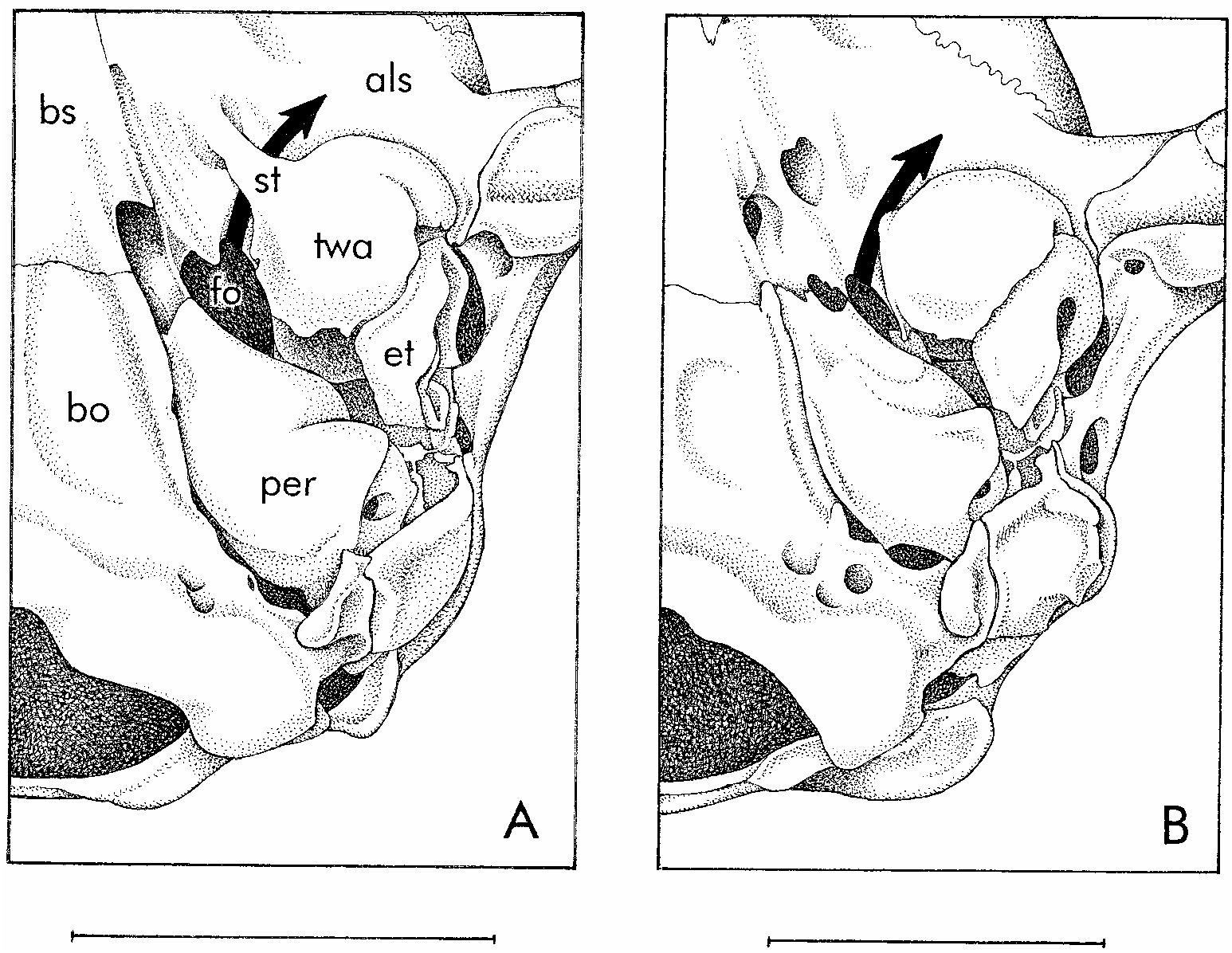
Close inspection of skulls revealed another difference between the two groups of specimens previously sorted by chromatic and morphometric traits. In specimens with fuscous dorsal fur, taller canines, and broader skulls, the lacrimal bone forms a prominent part of the anteroventral margin of the orbit; the lacrimal foramina then perforate the orbital margin, where they are exposed in lateral view (fig. 25B). By contrast, the lacrimal bone is not a prominent part of the anteroventral orbital margin in specimens with reddish dorsal fur, smaller teeth, and narrower skulls; in these, the lacrimal foramina are always located within the orbit, where they are moreorless concealed from lateral view (fig. 25A).
We interpret the correlation between pelage characters, morphometric differences, and lacrimal morphology described above as evidence that two species of Marmosops are sympatric at Paracou. Based on our examination of relevant type material (see below), we identify the reddish individuals with shorter canines, narrower skulls, and reduced lacrimals as M. parvidens , and the fuscous individuals with taller canines, broader skulls, and prominent lacrimals as M. pinheiroi . A single Paracou specimen (AMNH 267358), consisting of the skeleton only of a juvenile animal with a smashed skull, is not assignable with certainty to either species.
Marmosops parvidens (Tate) Figures 15B View Fig , 22A, 22B View Fig , 23A View Fig , 25A View Fig , 26A View Fig
VOUCHER MATERIAL: AMNH 266425 , 266426 , 267344 , 267347 , 267348 , 267350 , 267353 , 267359 , 267361 , 267817 ; MNHN 1995.927 – 1995.930 , 1995.933 . Total = 15 specimens .
IDENTIFICATION: Specimens of Marmosops parvidens can be distinguished from sympatric examples of M. pinheiroi at Paracou by their warmer (more reddish) dorsal fur, broader extent of selfcolored fur (which is never discontinuous between chin and anus), smaller teeth (especially upper canines), narrower skull, and by the reduced lacrimal contribution to the anteroventral orbital margin (see above). Because the difference in dorsal pelage color is subtle, and because there is some overlapping variation in the extent of selfcolored ventral fur, unambiguous species
TABLE 8 Principal Components Analysis of Craniodental Measurement Variation among 16 Adult Male Marmosops from Paracoua
identifications require cleaned cranial material. In our voucher series, height of canine (HC) affords the clearest discrimination (tables 9, 10), but toothwear and sexual dimorphism must be taken into account in sorting specimens by this criterion. Lacrimal morphology (fig. 25) is perhaps the most reliable cranial character for identifying juveniles with incompletely erupted (and therefore unmeasurable) canines.
Although a revision of what may be called the parvidens group of Marmosops is beyond the scope of this faunal report, we note that our conclusion that two species assignable to this complex are sympatric at Paracou is consistent with recent molecular evidence that M. parvidens sensu Pine (1981) is composite (Mustrangi and Patton, 1997; Patton et al., 2000). In addition to recognizing M. pinheiroi as a valid species, we note that M. juninensis (another ‘‘subspecies’’ of M. parvi dens sensu Pine, 1981) is equally distinctive and also merits specific recognition. Originally described by Tate (1931) on the basis of a single specimen (AMNH 63864), M. juninensis (now additionally represented by AMNH 230014–230016, all collected near Tarma in Depto. Junín, Peru) can be unambiguously distinguished from M. parvidens (sensu stricto) and M. pinheiroi by the size and shape of the male carpal tubercle (small er and not spoonshaped), by the consistent presence of posteromedial palatal vacuities, and by the absence of distinct accessory cusps on the upper canine (fig. 26). Like parvidens , but unlike pinheiroi , the lacrimal bone in juninensis does not form part of the anteroventral orbital margin, so the lacrimal foramen lies inside the orbit. Unlike any specimens of parvidens , however, the ventral pelage of juninensis is entirely graybased.
We examined the type of Marmosops parvidens bishopi (USNM 393535), the pelage of which Pine (1981) described as colored essentially like that of M. p. parvidens , but paler. In our opinion, this specimen represents another distinct species that differs from M. parvidens in lacking any trace of an anterior accessory cusp on the upper canine. Specimens from Bolivia (e.g., AMNH 268938) and Peru (e.g., AMNH 67243) with darker (more saturated) dorsal fur than the type may nevertheless be provisionally referred to M. bishopi based on upper canine morphology.
A specimen from northern Venezuela (USNM 371299) that Pine (1981) referred to Marmosops parvidens parvidens lacks an anterior accessory upper canine cusp (like M. bishopi ) and exhibits other differences from typical examples of M. parvidens as recognized in this report. The mystacial vibrissae of USNM 371299 appear to be very short, probably not extending much if at all beyond the base of the ear in life, whereas the mystacial hairs reach at least to the posterior margins of the pinnae in typical examples of M. parvidens . The dorsal fur of USNM 371299 is longer than in typical M. parvidens (about 9 mm middorsally versus about 7 mm), is fluffier in texture, and has a faintly marbled appearance that is not characteristic of other specimens that we refer to this species. Taken
TABLE 9
Measurements (mm) and Weights (g) of Adult Specimens of Marmosops parvidens
together, these differences suggest that USNM 371299 may represent an undescribed taxon, but more material should be examined to evaluate this conjecture.
With a single exception (AMNH 97333, see below), all of the specimens that we identify as Marmosops parvidens sensu stricto are from the Guiana subregion of Amazonia (fig. 27).
OTHER SPECIMENS EXAMINED: Brazil — Amazonas, Boca Rio Piratucu ( AMNH 93970 ), 80 km N Manaus ( USNM 579985– 579990 View Materials ) ; Para´, Ilha do Taiuna on lower Rio Tocantins ( AMNH 97333 ). French Guiana — Arataye ( USNM 548439 View Materials ) . Guyana — Demerara Mahaica, Hyde Park ( FMNH 18545 View Materials [holotype]) ; Upper Takutu Upper Essequibo, Karanambo ( ROM 97938) .
FIELD OBSERVATIONS: Because unvouchered sightings of Marmosops could not be unambiguously identified to species, all of our definite records of M. parvidens at Paracou are from collected specimens. Five specimens of M. parvidens were shot, one was caught by hand, one was caught in a Victor trap, one in a Sherman trap, and the rest (seven) were captured in pitfalls; all were collected at night. Nine specimens were taken on the ground, five were found perching in understory vegetation (usually on vertical stems) 0.2–1.5 m above the ground, and one was trapped on a liana 1.8 m above the ground. Seven specimens were collected in welldrained primary forest, five in creekside primary forest, one in swampy primary forest, one in primary forest of unspecified character, and one in secondary growth.
See the following account for habitat comparisons between Marmosops parvidens and M. pinheiroi .
Marmosops pinheiroi (Pine) Figures 16B View Fig , 19A View Fig , 20 View Fig , 21A View Fig , 22C, 22D View Fig , 23B View Fig , 25B View Fig
VOUCHER MATERIAL: AMNH 266423 , 266424 , 267007 , 267008 , 267341–267343 , 267345 , 267346 , 267349 , 267351 , 267352 , 267354 , 267357 ; MNHN 1995.925 , 1995.926 , 1995.931 , 1995.932 , 1995.934 . Total = 19 specimens .
IDENTIFICATION: See the preceding account for diagnostic comparisons with Marmosops parvidens .
In addition to the holotype and paratypes of Marmosops pinheiroi , we examined the type series of M. parvidens woodalli (USNM 393529–393532, 393534, 545543), a subspecies that Pine (1981) described from the vicinity of Belém, Brazil. Although these specimens average paler dorsally than examples of M. pinheiroi from north of the Amazon, they are otherwise similar in pelage and craniodental characters, and we provisionally regard them as conspecific. An adult male specimen from the right bank of the lower Rio Xingu (USNM 549294), however, may represent an undescribed taxon. Although most of its qualitative traits match those of M. pinheiroi , it is much paler dorsally and has substantially smaller upper canines (HC = 1.06 mm) for its sex than any example of that species as recognized by us.
OTHER SPECIMENS EXAMINED: Brazil — Amapa´ , Serra do Navio ( USNM 461459 View Materials [holotype], 461460, 461462–461465) ; Para´ , Belém ( USNM 545543 View Materials ), Utinga ( USNM 393529–393532 View Materials , 393534 View Materials ) . Guyana — Pota
TABLE 10 Measurements (mm) and Weights (g) of Adult Specimens of Marmosops pinheiroi
ro Siparuni, Iwokrama Reserve (ROM 108920). Venezuela — Bolívar, Auyantepui (AMNH 130521, 130568, 130570), Churitepui (AMNH 176352, 176353), 85 km SE El Dorado (USNM 385046).
FIELD OBSERVATIONS: All of our definite records of Marmosops pinheiroi at Paracou are from collected specimens. Four specimens were shot, three were captured in Victor traps, and the remainder (12 specimens) were caught in pitfalls; all were taken at night. Thirteen specimens were trapped or shot on the ground (one in a hollow log), but six were taken 0.3–1.5 m above the ground on vertical stems and lianas. Four specimens were collected in welldrained primary forest, five in creekside primary forest, six in swampy primary forest, and four in secondary growth.
Marmosops parvidens and M. pinheiroi clearly overlap in habitats at Paracou. Although our ecological classification is coarse and doubtless obscures many subtle differences among capture sites with the same descriptor (e.g., ‘‘welldrained primary for est’’), we sometimes took both species in close proximity. One line of pitfall traps that traversed 50 m of apparently homogeneous primary forest along a small stream, for example, captured three parvidens and four pinheiroi between 29 July and 13 August 1993. Nevertheless, it is noteworthy that whereas we rarely took parvidens in swampy forest or secondary growth, over half of our pinheiroi specimens were collected in those habitats. Both species appear to occur only in the forest understory: no Marmosops were sighted or trapped at heights greater than about 2 m above the ground.
None of the adult female Marmosops we collected were carrying suckling young.






| ROM |
Royal Ontario Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |